乳品化学

乳中主要成份为水、脂肪、蛋白质、乳糖和矿物质(无机盐类),乳中还含有其它微量成份,例如:色素、酶类、维生素和磷脂(具有类脂特性)以及气体。
乳是哺乳动物乳腺分泌的一种不透明白色液体,主要功能是哺育新生幼体,确保生长发育所必需营养素的摄入。因此,其化学成分极为复杂,且可能受多种因素影响,包括气候条件、泌乳阶段、牲畜年龄、健康状况、日粮配置以及品种差异。
乳中主要成份为水、脂肪、蛋白质、乳糖和矿物质(无机盐类),同时还含有其它微量成份,例如:色素、酶类、维生素和磷脂(具有类脂特性),以及气体。除去乳中的水份和气体,剩下的物质我们称之为干物质(DM)或乳的总固形物。
为阐述乳中各种成份及其如何受乳品加工各阶段处理工艺的影响,有必要运用化学术语进行表述,因此,本章乳品化学将从简要回顾一些基本化学概念入手。
C 碳
Ca 钙
Cl 氯
H 氢
I 碘
K 钾
N 氮
Na 钠
O 氧
P 磷
S 硫
基本化学概念
物质由原子构成,原子结合在一起形成物质和化合物。物质的复杂性取决于原子和分子的组织方式及相互作用,从而形成具有独特结构和性质的各类物质与化合物。
原子
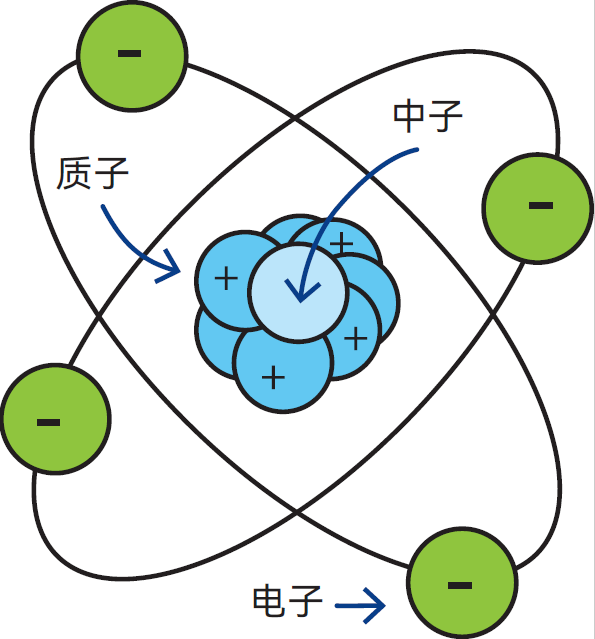
图2.1 原子核由质子和中子组成,电子绕原子核运动
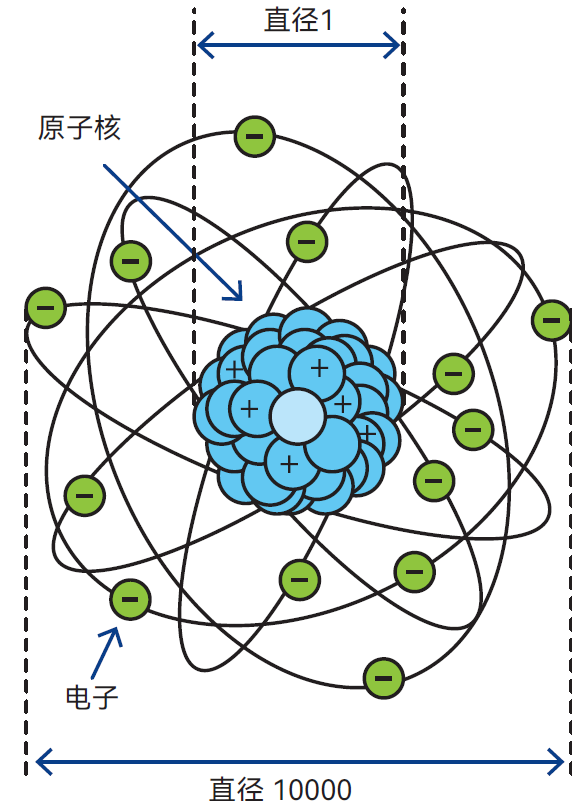
图2.2 与原子相比,原子核实在太小,如果将其放大成一个网球大小,则最外电子壳层将离中心325米
原子是构成自然界所有物质的最小单元,无法通过化学方式再分割。由同类原子组成的物质称为元素。目前已知元素有100多种,如氧、碳、铜、氢和铁。然而,自然界中大多数物质由多种不同元素组合而成。例如:空气是氧气、氮气、二氧化碳和其它稀有气体的混合物,而水则是由氢元素与氧元素组成的化合物。
原子核由质子和中子组成(图2.1)。质子带一个正电荷,而中子呈电中性。绕核运动的电子所带负电荷与质子的单位正电荷电量相等但电性相反。
由于原子包含的质子与电子数量相等,且正负电荷数相同,因此原子整体呈电中性。
原子非常小(图2.2)。一枚小铜币中所含原子数相当于一千兆年中所含秒数!尽管如此,原子内部绝大部分仍是空无一物的空间。若将原子核的直径设定为1,则整个原子的直径约为10000。
离子
一个原子可能失掉或得到一个或多个电子,此类原子不再保持电中性,被称为离子。若离子所含电子数多于质子,则带负电;若失去一个或多个电子,则带正电。
正离子与负离子总是同时存在,即在溶液中以阳离子(带正电)和阴离子(带负电)形式存在,或以固态盐的形式存在。普通食盐由钠离子(Na+)和氯离子(Cl-)构成,其化学式为NaCl(氯化钠)。
分子
同种或不同种元素的原子可结合形成更大的单元,称为分子。分子能进一步构成固态物质(如铁(Fe)和二氧化硅(SiO2))、液体(如水(H2O)),或气体(如氢气(H2))。若分子主要由碳(C)原子、氢(H)原子、氧(O)原子构成,则形成的化合物称为有机物,即由有机元素生成。以乳酸(C3H6O3)
为例:其分子式表明,该分子由3个碳原子、6个氢原子和3个氧原子组成。
分子中的原子数可能相差很大,有的分子只有两个原子相连而成,有的分子则由数百个原子组成。
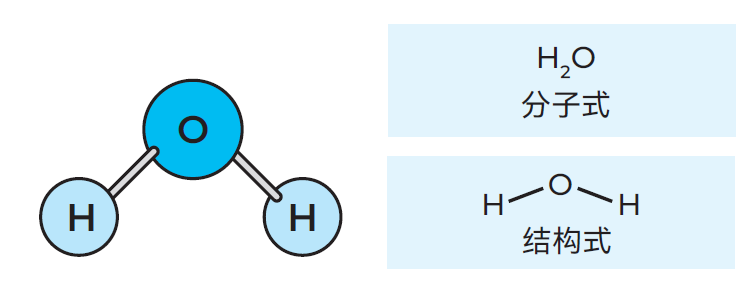
图2.3 一个水分子的三种表达方式
牛奶的基本理化特性
牛奶是一种复杂的多分散体系,由约87%的水和13%的干物质组成(表2.1)。这些干物质悬浮或溶解在水中,根据固体类型、粒径及各组分间亲和力的不同(表2.2),它们在水相中可形成多种分散系(混合体系)(图2.4)。
参考资料:《乳业词典》(A Dictionary of Dairying),作者:J G Davis
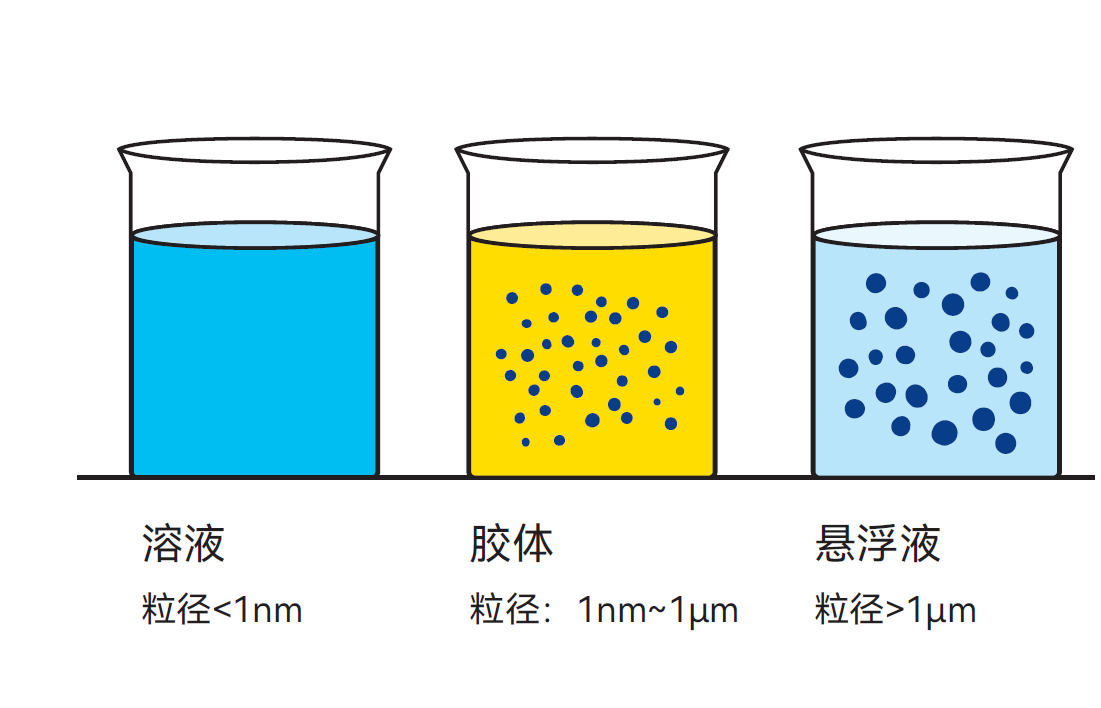
图2.4 水相中的多种分散系
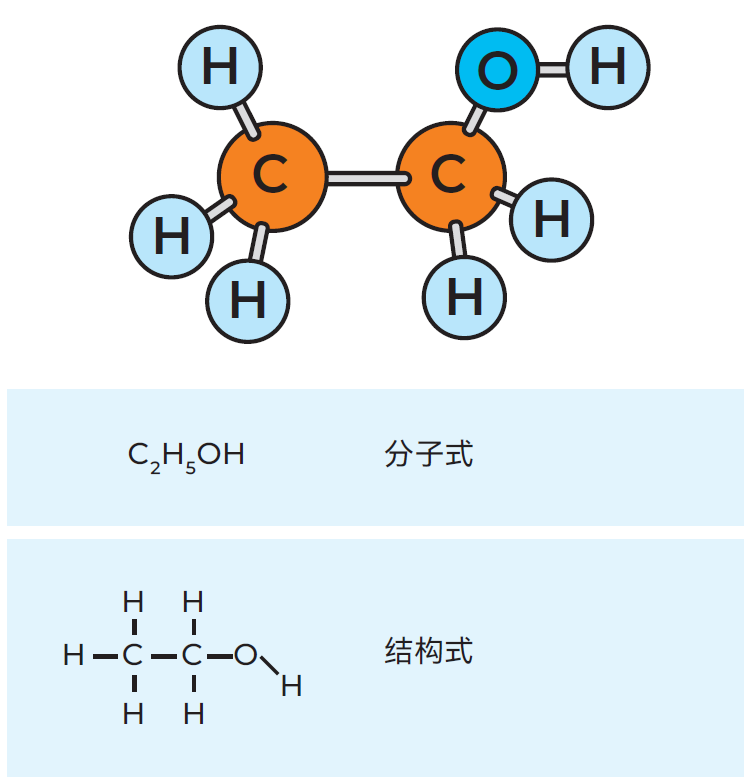
图2.5 一个乙醇分子的三种表达方式
定义
溶液: 指次要成分(溶质)均匀分散于主要成分(溶剂)中形成的液态混合物。例如,盐溶解在水中形成盐水溶液。
悬浮液: 指固体微粒分散于液相中形成的异质混合物,即使由微小固体颗粒构成,这些固体也不能溶解,只能保持悬浮状态。
乳液: 一种液体的液滴悬浮于另一种液体中形成的悬浮液。牛奶属于水包油型乳液,黄油则为油包水型乳液(图2.6)。被分散的液体称为分散相,另一个称为连续相。
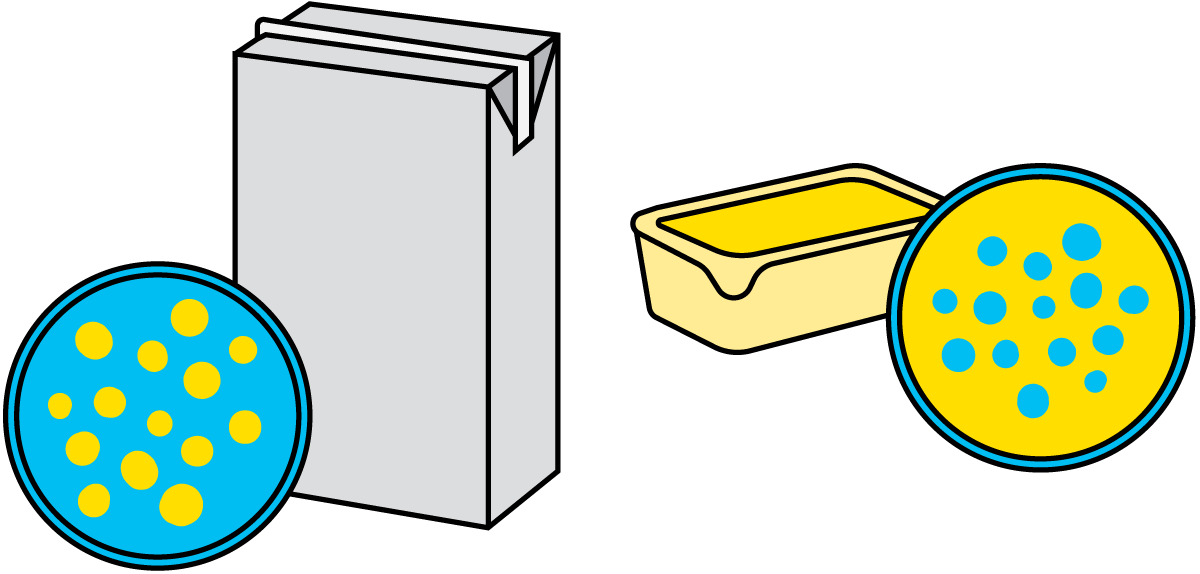
图2.6 牛奶中的水包油型乳液和与黄油中的油包水型乳液
• 微小粒径
• 携带电荷
• 颗粒对水分子具有亲和力
胶体溶液: 当物质以介于真溶液(如糖水)和悬浮液(如粉笔水)之间的分散状态存在时,称为胶体溶液或胶体悬浮液。
胶体的典型特征包括:
- 微小粒径
- 携带电荷
- 颗粒对水分子具有亲和力
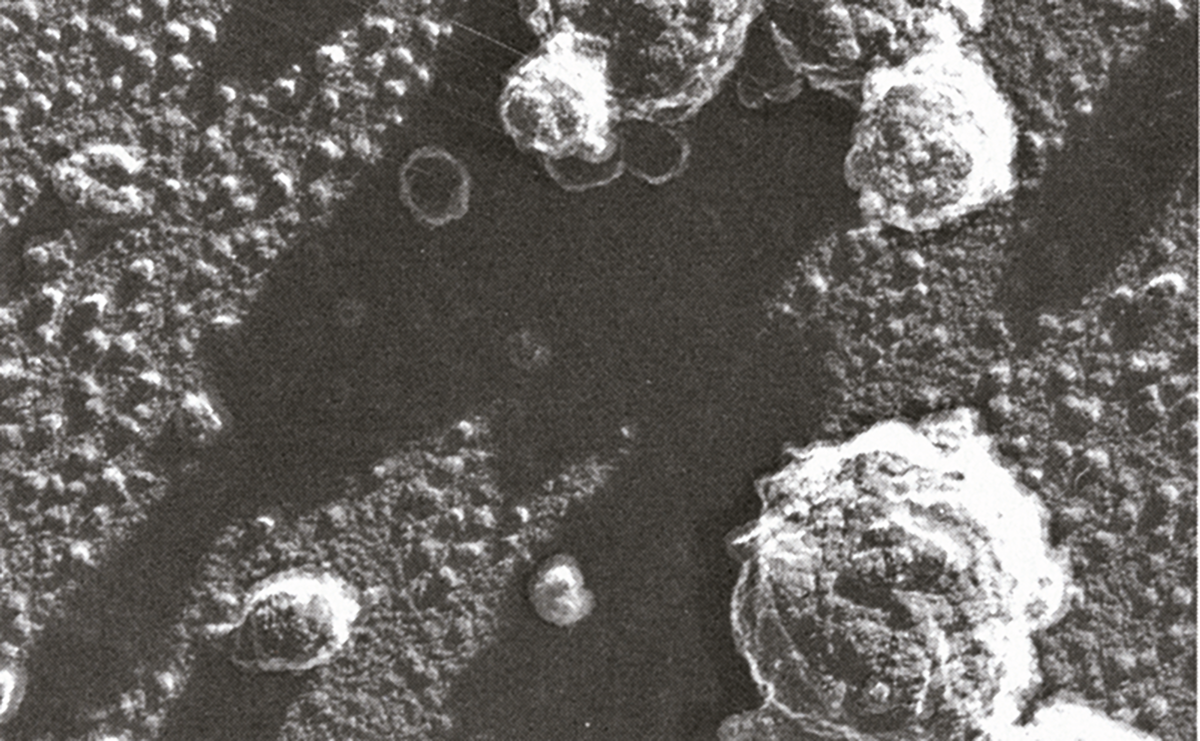
图2.7 通过电子显微镜可观察到乳蛋白。
参考资料:Miloslav Kaláb,《食品结构》,1993年
盐类等物质可通过改变水合作用来破坏胶体系统的稳定性,从而降低蛋白质的溶解度。热量等因素会导致乳清蛋白结构伸展并增加蛋白质间相互作用,酒精则可能使蛋白质颗粒脱水。
真溶液:物质与水或其他液体混合即形成真溶液。真溶液可分为:
- 非离子溶液:当乳糖溶于水时,其分子结构无重大变化。
- 离子溶液:当食盐溶解于水中时,阳离子(Na+)和阴离子(Cl-)分散在水中,形成电解液(图2.8)。
溶液的酸碱度
当一种酸(例如盐酸,HCl)与水混合时,会释放出带正电荷(H+)的氢离子(质子)。这些氢离子迅速与水分子结合,形成水合氢离子(H3O+)。把碱(金属氧化物或氢氧化物)加入水中时,可形成碱性溶液。碱溶解时会释放出氢氧根离子(OH-)。
- 若溶液中氢氧根离子与氢离子数量相等,则该溶液呈中性(图2.9)。
- 若氢氧根离子多于氢离子,则溶液呈碱性(图2.10)。
- 若氢离子多于氢氧根离子,则溶液呈酸性(图2.11)。
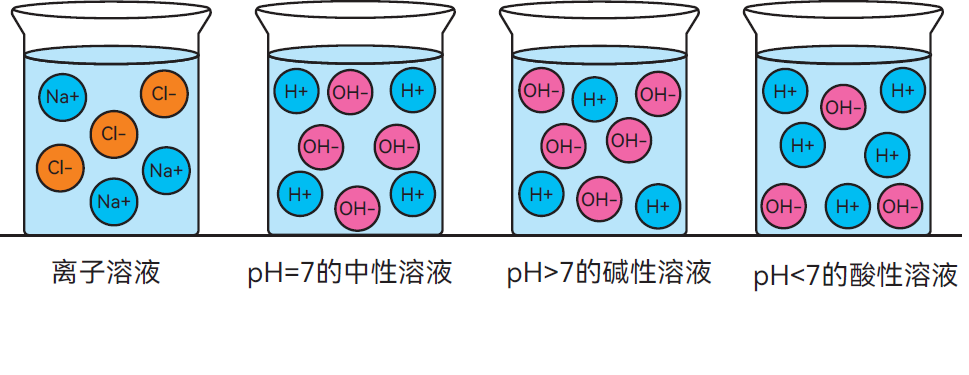
图 2.7-2.11
pH值
溶液的酸度由氢离子浓度决定,但不同溶液间的氢离子浓度差异显著。pH符号用于表示氢离子浓度。
其数学定义为氢离子摩尔浓度的负对数(以10为底),即: pH = – log [H+].
由此在25℃时得出以下标度:
pH = 7 中性溶液
pH < 7 酸性溶液
中和反应
当酸与碱混合时,氢离子与氢氧根离子相互反应生成水。若按特定比例混合酸和碱,所得混合物将呈中性——既无过量氢离子也无过量氢氧根离子,pH值为7。此过程称为中和反应,其化学反应式为:
中和反应会生成盐。当盐酸(HCI)与氢氧化钠(NaOH)溶液混合时,两者发生反应,生成氯化钠(NaCl)和水(H2O)。由盐酸形成的盐类称为氯化物,其它盐类也同样以参与反应的酸类命名,如柠檬酸形成柠檬酸盐,硝酸生成硝酸盐,依此类推。
扩散
溶液中的粒子(离子、分子或胶体)在力的作用下,从高浓度区域向低浓度区域移动的过程,称为扩散。扩散过程持续进行,直至整个溶液浓度均匀一致。糖在咖啡中的溶解,就是典型的扩散现象。糖在热饮中快速溶解,糖分子通过扩散作用均匀分布至整个饮品中。
扩散速率取决于颗粒速度,而颗粒速度又取决于温度、粒径及溶液各部分之间的浓度差。
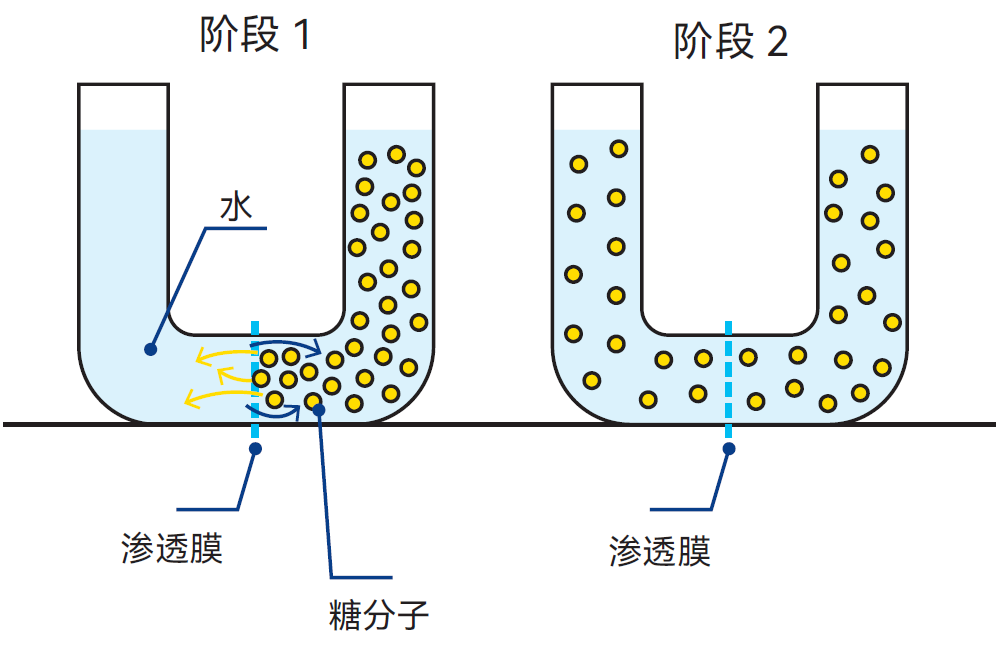
图2.12 通过渗透膜,糖分子与水分子以相反方向扩散,最终实现溶液浓度平衡
图2.12展示了扩散过程的原理。U形管被渗透膜分成两个部分左侧注入水,右侧注入可透过渗透膜的糖溶液。经过一段时间的扩散后,膜两侧的糖浓度趋于相等。
渗透
渗透是指透过适宜的膜,纯水自发流入水溶液,或低浓度溶液自发流向高浓度溶液的现象。渗透现象可通过图2.13中示例来说明:U形管被半渗透膜分成两个部分。左侧注入水,右侧注入不能透过渗透膜的糖溶液。此时水分子会透过膜扩散至糖溶液中,并将其稀释至较低浓度,此过程即称为渗透。
糖溶液稀释后体积增加,液面上升,如图2.13所示,溶液作用于膜上的静压a大于另一侧水的静压。在此压力失衡状态下,水分子在溶液侧较高静压作用下开始反向扩散。
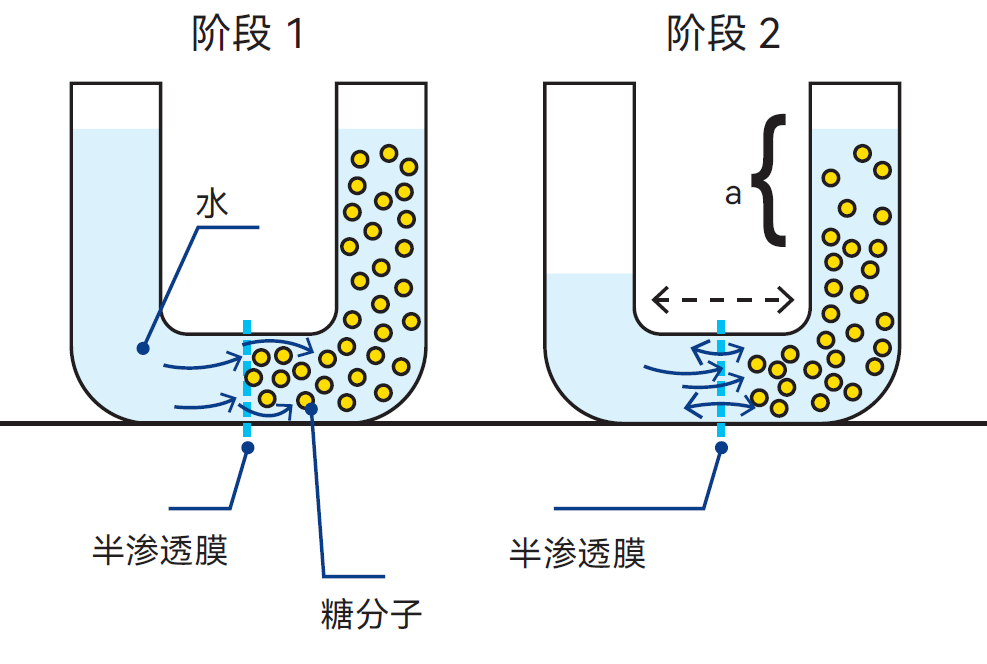
图2.13 糖分子太大,无法透过半渗透膜扩散,仅有体积较小的水分子能够扩散,达到浓度平衡。“a”为溶液的渗透压
当双向水扩散速率相等时,系统达到平衡。若一开始就对糖溶液施加静压,则穿过渗透膜进入糖溶液的水分子将会减少。为阻止水分子扩散到糖溶液中,避免达到浓度均衡,所需施加的流体静压,称为该溶液的渗透压。
反渗透
若对糖溶液施加高于渗透压的压力,可促使水分子从糖溶液向水中扩散,进而使糖溶液浓度升高。该过程如图2.14所示,在商业上用于浓缩溶液,称为反渗透(RO)。
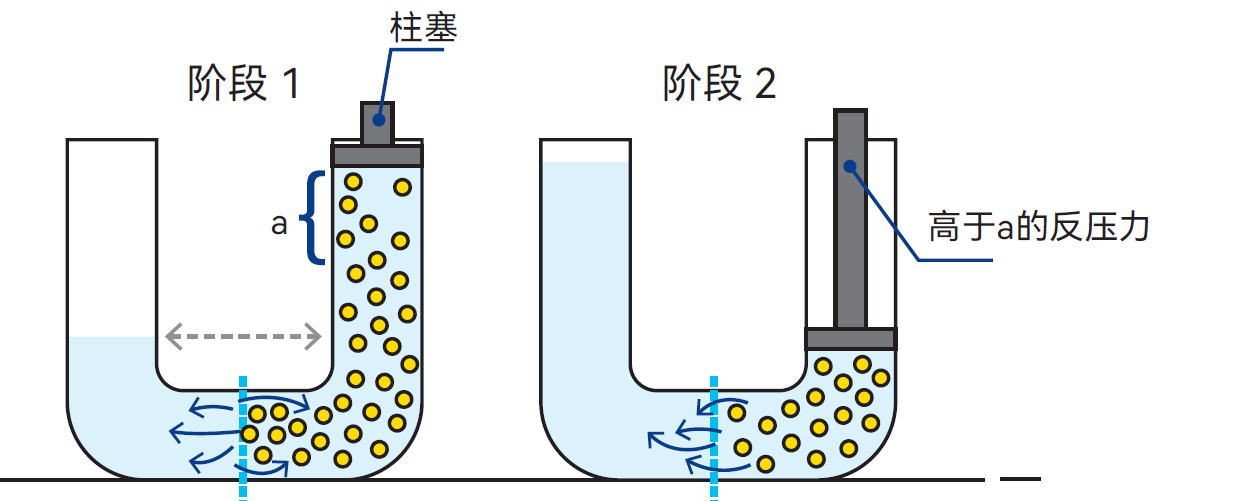
图2.14 若对糖溶液施加高于渗透压的压力,水分子扩散, 溶液浓度升高
渗析
渗析是借助浓度差为驱动力,把溶液中的大颗粒与小颗粒分离的技术。例如:将蛋白质与盐类分离。将待处理溶液置于膜的一侧,溶剂(水)置于另一侧。体积较小的盐分子可通过膜的孔径,但蛋白质分子则无法通过(见图2.15)。扩散速率随浓度差而变化,因此经常更换膜另一侧的溶剂可加速渗析过程。
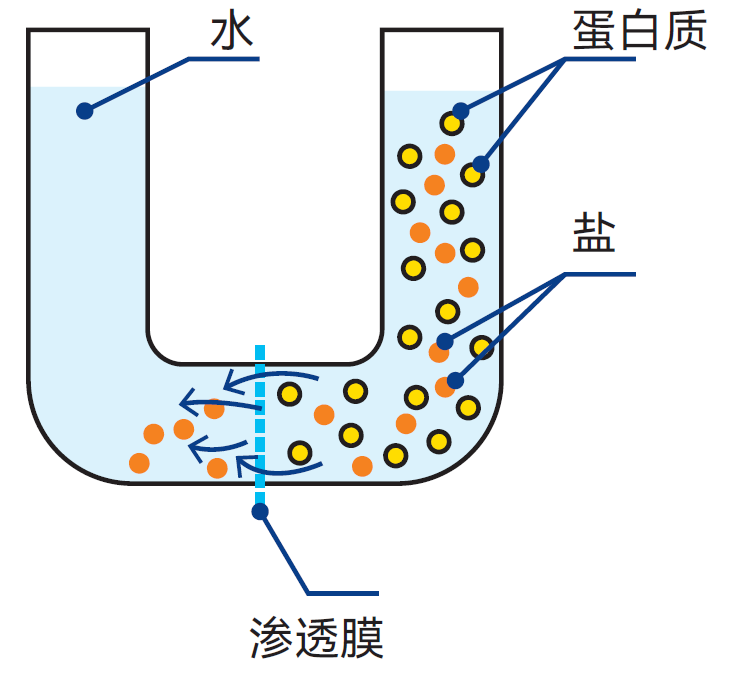
图2.15 在膜的一侧稀释溶液,当小分子穿过膜时,大分子的浓度会相应升高
牛奶的成分组成
不同品种奶牛及同一品种个体间,牛奶中各主要成分的含量可能存在显著差异。因此,仅能提供含量变化的限值。表2.3中数据仅为示例性数值。
除总固形物之外,非脂乳固体(SNF)这一术语也用于描述牛奶成分。SNF即除脂肪外的总固形物含量。根据表2.3中所提供的数据,SNF的平均含量为13.0%-3.9%=9.1%。常乳的pH值通常在6.6-6.8之间,最常见平均值约为6.7。相应的测定温度约为25℃。
乳脂肪
牛奶和奶油是典型的水包脂型(或水包油型)乳液。乳脂肪以小球或液滴形态分散于乳浆中(见图2.16)。其直径范围为0.1-20μm(1μm=0.001mm),平均直径为3-4μm,每毫升牛奶中约含1010个脂肪球。
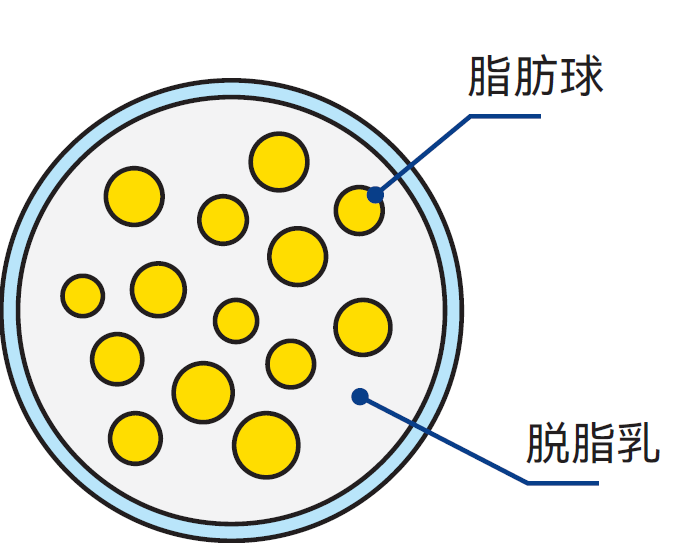
图2.16 牛奶微观结构示意图
该乳液由一层极薄的膜结构维持稳定。此膜厚度仅为10-20nm(1nm=10-9m),包裹于脂肪球外且成分复杂。乳脂肪由甘油三酯(主要组分)、甘油二酯与单酯、脂肪酸、固醇、类胡萝卜素(赋予脂肪黄色色泽)和维生素(A、D、E、K)构成。微量元素为次要组分。乳脂肪球的组成如图2.17所示。
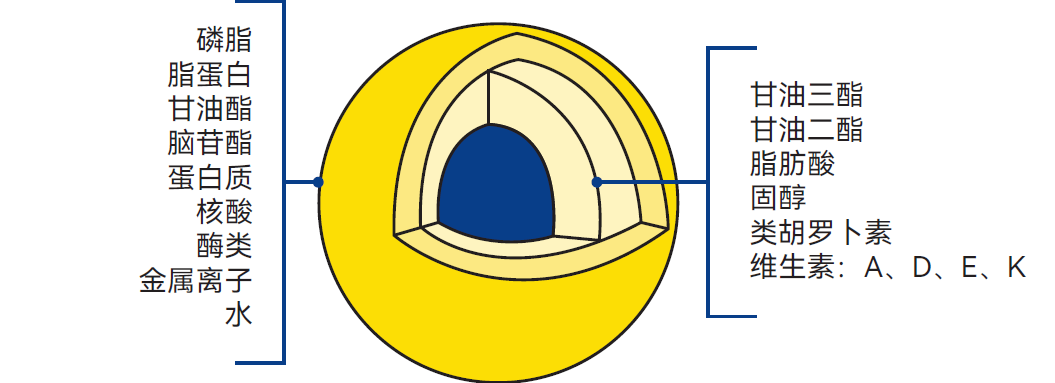
图2.17 乳脂肪的组成
球径为0.1-20μm,平均球径为 3-4μm。
膜由磷脂、脂蛋白、脑苷脂、蛋白质、核酸、酶类、微量元素(金属离子)和结合水组成。应注意的是,膜的组成与厚度并非恒定,因其成分会与周围乳浆持续进行动态交换。
由于脂肪球不仅是牛奶中最大的颗粒,同时也是最轻的颗粒(15.5℃时密度=0.93g/cm3),因此当牛奶静置存放时,脂肪球会自然上浮至牛奶的表层(见图2.18)。
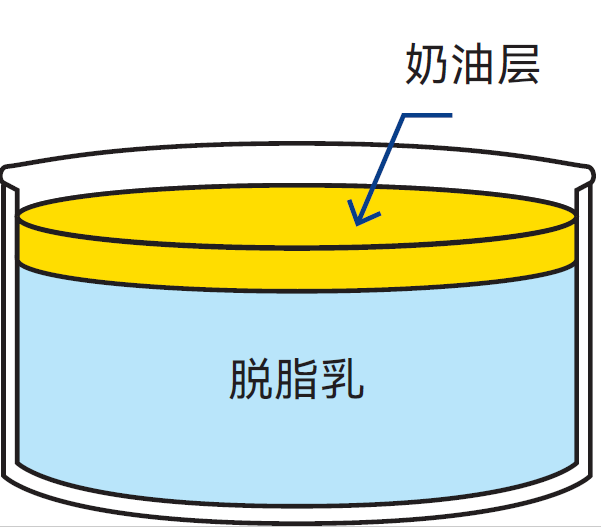
图2.18 如果将牛奶静置,脂肪将会上浮并在表层形成奶油层
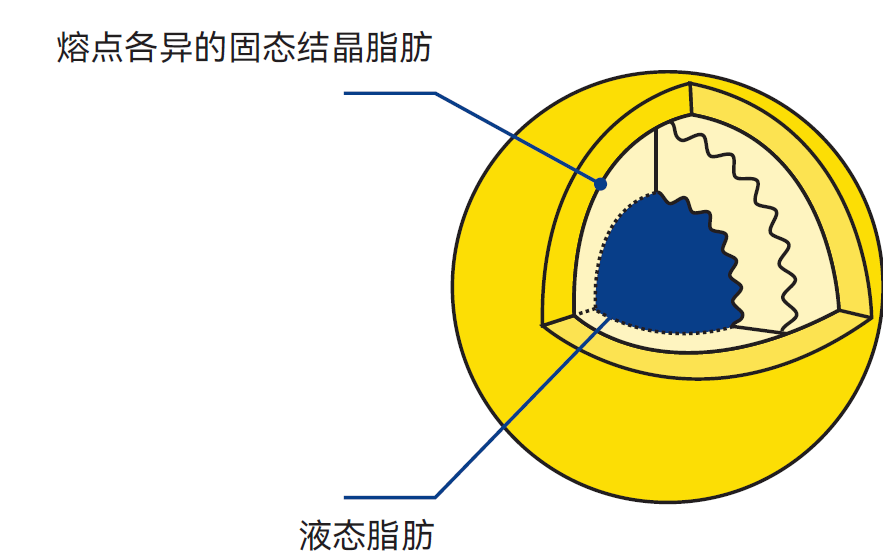
图2.19 脂肪球剖视图
脂肪球上浮速率遵循斯托克斯定律,但其微小尺寸导致自然脂肪上浮的过程极为缓慢。然而,在一种名为凝集素的蛋白质作用下,脂肪球会发生聚集,进而加速乳油分离。这些聚合体的上浮速率远高于单个脂肪球。此类聚合体易被加热或机械处理破坏。例如,在75℃/2分钟的时间-温度条件下,凝集素会发生变性,脂肪球聚集能力也随之消失。
乳脂肪的化学结构
当乳从乳腺排出时,乳脂肪呈液态(37℃),这意味着脂肪球在历经中度的机械处理(例如在管道中泵送、流动)时,可轻松改变自身形状,且不会从其膜结构中脱离。
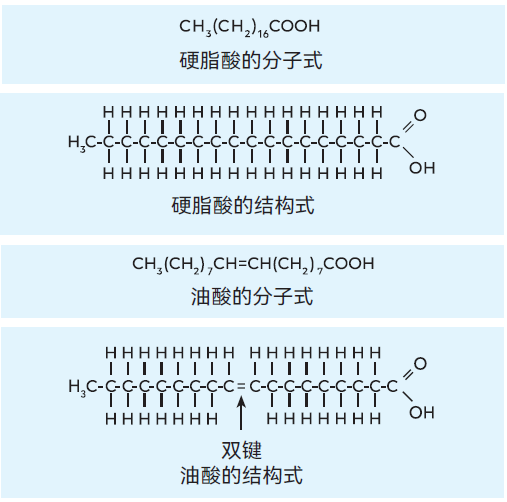
图2.20 硬脂酸和油酸的分子式和结构式
所有脂肪均属于一组名为酯的化学物质,酯是醇与酸反应形成的化合物。乳脂肪是不同脂肪酸酯(名为甘油三酯)的混合物,甘油三酯由一种名为甘油的醇类物质和多种脂肪酸组成。甘油酯约占乳脂肪总量的99%。
脂肪酸分子由一个烃链和一个羧基组成(分子式为RCOOH)。
在饱和脂肪酸分子中,碳原子通过单键连接成链,而在不饱和脂肪酸分子中,烃链上有一个或多个双键,见图2.20。每个甘油分子可结合三个脂肪酸分子,且三者种类可不同,因此乳脂肪中甘油酯的种类极为丰富。
表2.4列出了乳脂肪甘油三酯中最重要的脂肪酸。
脂肪熔点
如表2.4所示,牛奶中含量最丰富的四种脂肪酸分别是肉豆蔻酸、棕榈酸、硬脂酸和油酸。在室温下,前三种脂肪酸呈固态,后者呈液态。如引用数据所示,不同脂肪酸的相对含量可能存在显著差异,而这种差异会影响脂肪的硬度。高熔点脂肪酸(如棕榈酸)含量高的脂肪质地坚硬;反之,低熔点油酸含量高的脂肪则产出软质黄油。测定每种脂肪酸的具体含量属于科研范畴,在实际应用中,只需确定一个或几个能反映乳脂肪结构特征的常数或指数即可。
碘值
碳氢原子数相同但单双键数不同的脂肪酸具有截然不同的特性。表征其特性的最重要、最广泛应用的方法是测定脂肪的碘值(IV)。碘值指脂肪所能结合的碘的百分比。碘会被不饱和脂肪酸的双键所吸收。由于油酸是目前不饱和脂肪酸中含量最丰富的一种,室温下呈液态,因此碘值主要是油酸含量的衡量指标,也是脂肪软硬程度的衡量指标。
乳脂的碘值通常在24~26之间,其波动取决于奶牛饲料构成:夏季的绿色牧草促进油酸生成,故而夏季的乳脂肪柔软(碘值高)。葵花籽饼和亚麻籽饼等精饲料也能产生软质脂肪,而椰子油饼、棕榈油饼和根茎类蔬菜的茎叶则能产生硬质脂肪。因此通过选择适宜日粮可调控乳脂稠度。
图2.21展示了瑞典地区乳脂碘值在一年内的变化。
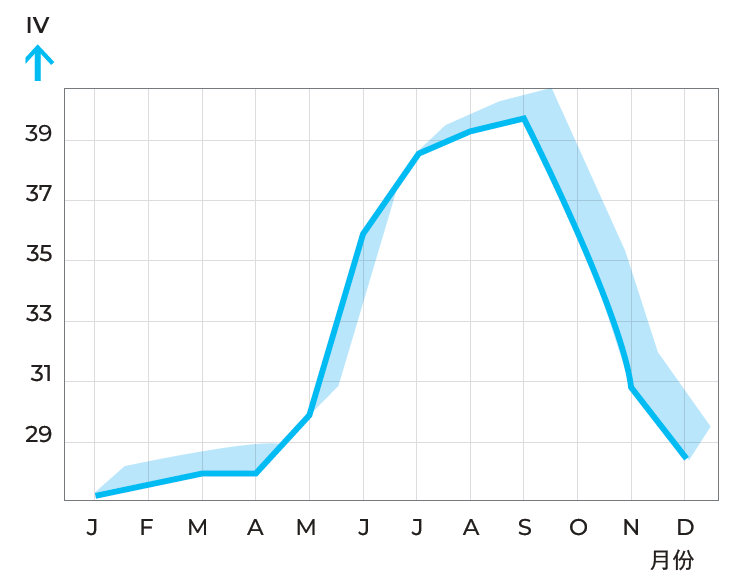
图2.21 一年中不同时期碘值变化,碘值是衡量脂肪中油酸含量的指标
折射率
脂肪中不同脂肪酸的含量同样会影响其折射光线的方式。因此,测定脂肪的折射率并据此计算碘值已成为常规实践,这是一种快速评估脂肪硬度的方法。
核磁共振(NMR)
除测定碘值或折射率外,还可通过脉冲核磁共振(NMR)直接测定饱和脂肪与不饱和脂肪的比率。若需转换,可通过换算因子将NMR测定值转换为相应碘值。NMR法还可用于探究脂肪结晶程度与结晶时间的函数关系。研究表明,将40%脂肪含量的奶油从60℃冷却至5℃后,脂肪结晶过程较为缓慢——至少需要2小时才能使结晶脂肪占比达到总量的65%,见图2.22。
同时还发现,温度降至5℃两分钟后,结晶脂肪比例仅为15-20%。乳脂的NMR值通常在30~41之间。
脂肪结晶
结晶过程中的脂肪球处于高度敏感状态,即使只是经受中度的机械处理,也极易受损破裂。
电子显微镜研究表明,脂肪以单分子球体形式结晶,如图2.22所示。与此同时,脂肪发生分馏,熔点最高的甘油三酯形成外层球体。由于结晶脂肪比容小于液态脂肪,球体内会产生张力,导致结晶期脂肪球结构极不稳定、易破裂。其结果是液态脂肪球被释放到乳浆中,游离脂肪作为粘合剂使未破裂脂肪球聚集成块(与黄油制作中的现象相同)。脂肪结晶会释放熔化热,导致温度小幅上升(40%含脂率的奶油从60℃冷却至7~8℃后,结晶期间温度会回升3~4℃)。
在生产各类奶油制品时,必须牢记乳脂的这一重要特性。
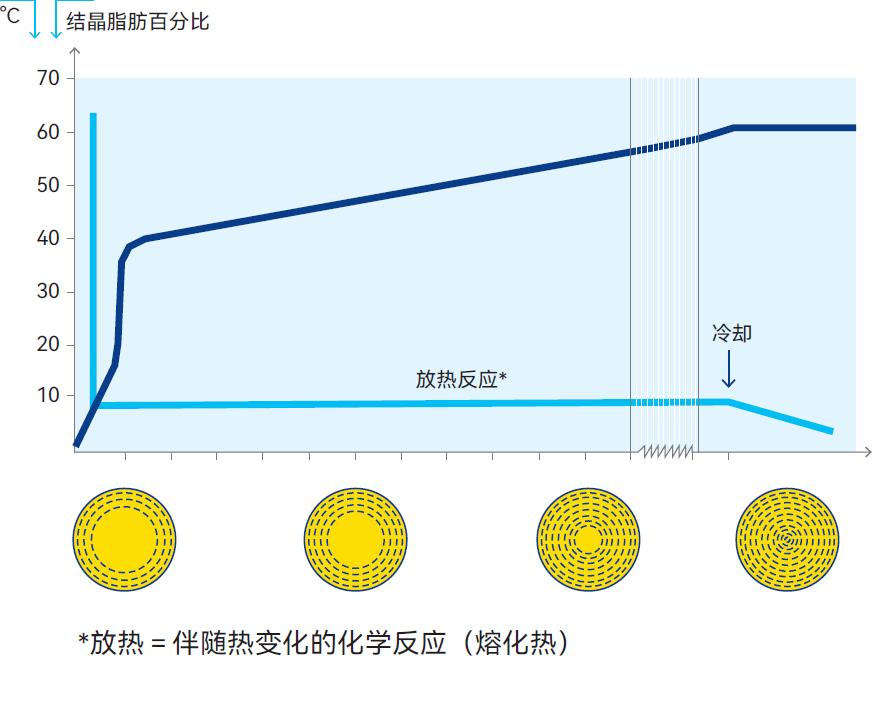
图2.22 乳脂肪结晶是一种放热反应,即该化学反应伴随热量释放。本结晶曲线基于NMR法分析绘制
牛奶中的蛋白质
蛋白质是人类膳食的重要组成部分。我们摄入的蛋白质经肝脏和消化系统的作用,被分解为较简单的化合物,随后输送至全身各细胞,成为构建人体自身蛋白质的材料。生物体内绝大多数化学反应由特定活性蛋白质(酶)所调控。
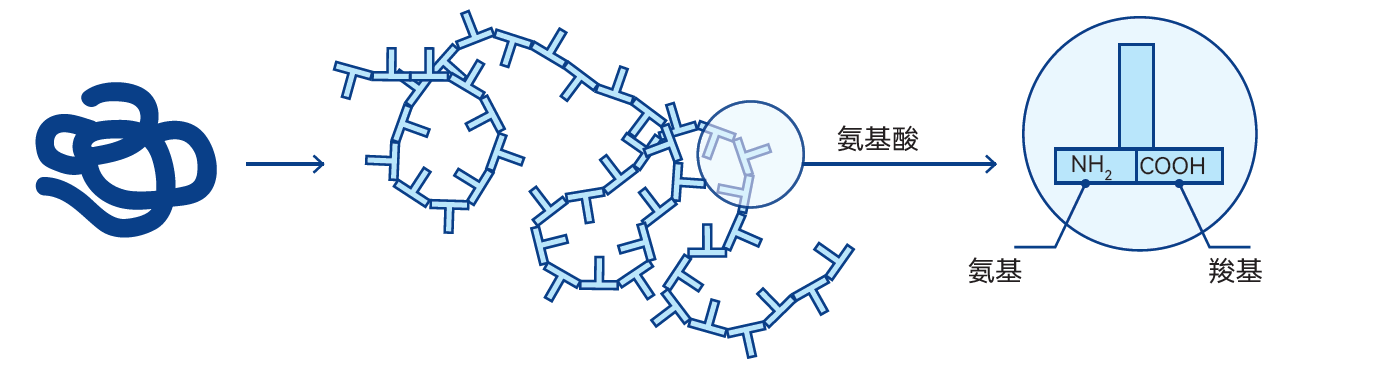
图2.23 由氨基酸、氨基和羧基组成的蛋白质分子链模型
蛋白质是由名为氨基酸的小分子单元构成的大分子物质(图2.23)。蛋白质分子由一条或多条氨基酸链通过特定顺序连接而成,通常包含100-200个氨基酸,但也存在由更少或更多氨基酸构成的蛋白质分子。
氨基酸
T图2.24中的氨基酸是构成蛋白质的基本单元,其分子的显著特征是同时含有一个氨基(-NH2)和一个羧基(-COOH)。蛋白质由一种特定氨基酸(α-氨基酸)构成,即氨基与羧基均结合在同一个碳原子(α-碳原子)上的氨基酸。氨基酸属于一类能在碱性溶液中释放氢离子和在酸性溶液中吸收氢离子的化合物。
此类化合物称为两性电解质或两性物。因此氨基酸可呈现三种状态:
- 在碱性溶液中带负电荷
- 正负电荷数量相等时呈中性
- 在酸性溶液中带正电荷
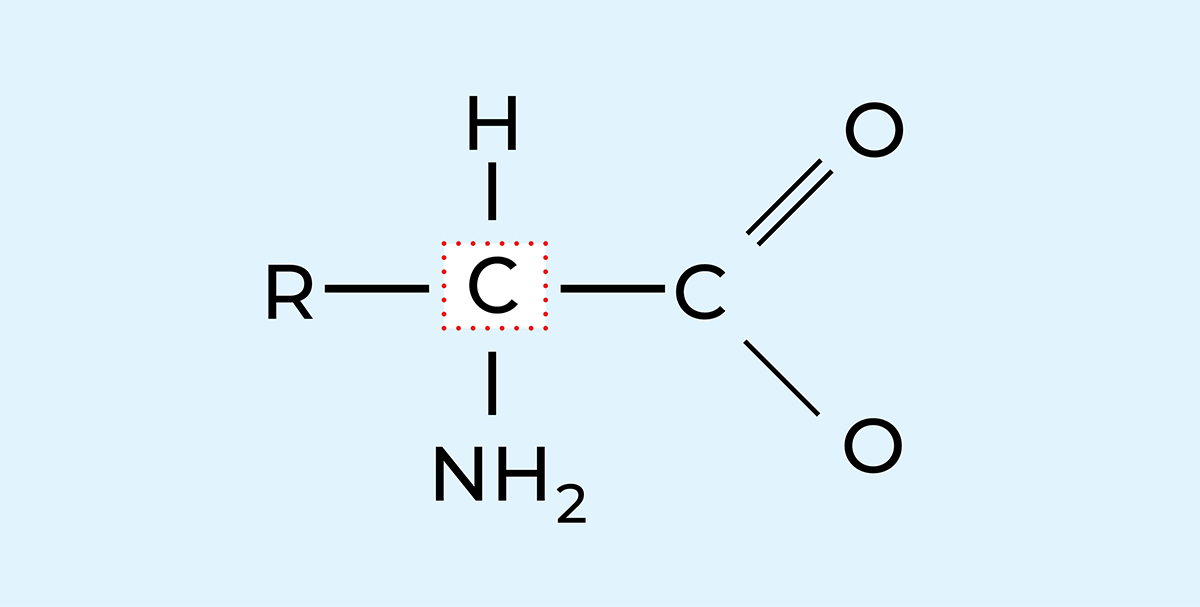
图2.24 一般氨基酸的结构式,图中R代表与中心碳原子结合的有机物
蛋白质由20种氨基酸组成。从营养学角度来说,一个重要的事实是:在这20种氨基酸中,有8种(婴儿为9种)无法由人体自身合成。由于这些氨基酸对维持正常的新陈代谢至关重要,因此必须通过食物摄取。这些氨基酸被称为必需氨基酸,且全部存在于乳蛋白中。
乳蛋白分子中氨基酸的类型及排列顺序决定了蛋白质的特性。分子链中氨基酸种类或位置的任何改变,都可能产生具有不同性质的蛋白质。
由于在一条含有100-200个氨基酸的链中,20种氨基酸的可能组合方式极多,因此特性各异的蛋白质数量也非常可观。图2.24是一个氨基酸模型。如前所述,氨基酸同时含有弱碱性氨基(-NH2)和弱酸性羧基(-COOH)。这些基团与一个侧链(R)共同连接在中心碳原子上。
若侧链为极性基团,则除极性侧链外,碱性基团和酸性基团的吸水性通常会占主导地位,使整个氨基酸亲水且易溶于水。这种氨基酸被命名为亲水性(爱水)氨基酸。
另一方面,如果侧链是不含亲水基团的烃链,则烃链的特性将占主导地位。长烃链会排斥水,使氨基酸的溶解性或与水的相容性降低。这种氨基酸称作疏水性(憎水)氨基酸。
若疏水烃链中存在羟基( -OH) 或氨基(-NH2)等某些自由基,可减弱疏水性并向亲水性转化。如果疏水性氨基酸在蛋白质分子的一部分中占主导地位,则该部分将具有疏水特性。
亲水性氨基酸在分子另一部分聚集,同样会赋予该部分亲水性。因此蛋白质可表现为亲水、疏水、介于亲水与疏水之间,或局部亲疏水特性。
部分乳蛋白分子内部的水相容性存在显著差异,而蛋白质的诸多重要特性正取决于这些差异。
酪蛋白中某些氨基酸链的羟基可与磷酸发生酯化。这些基团使酪蛋白能结合钙离子或胶体羟基磷酸钙,在分子之间或单个分子内形成牢固的桥接。
乳蛋白的带电状态
乳蛋白中部分氨基酸侧链携带电荷,其带电状态由牛奶的pH值决定。当通过加酸或加碱改变牛奶的pH值时,蛋白质的电荷分布也将发生改变。乳蛋白的带电状态以及由此产生的特性如图2.25~2.28所示。
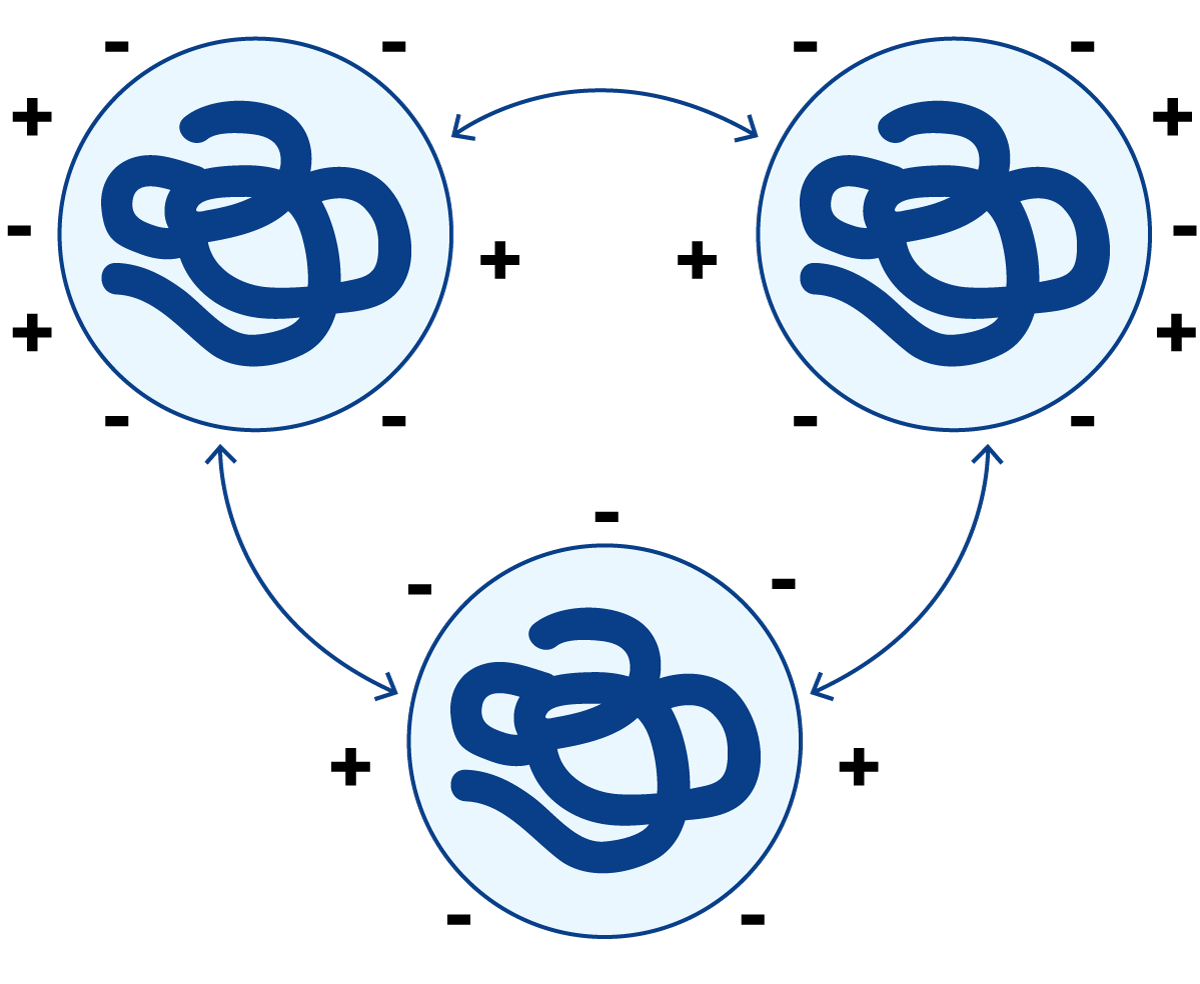
图2.25 pH=6.6时,蛋白质分子所带净电荷为负
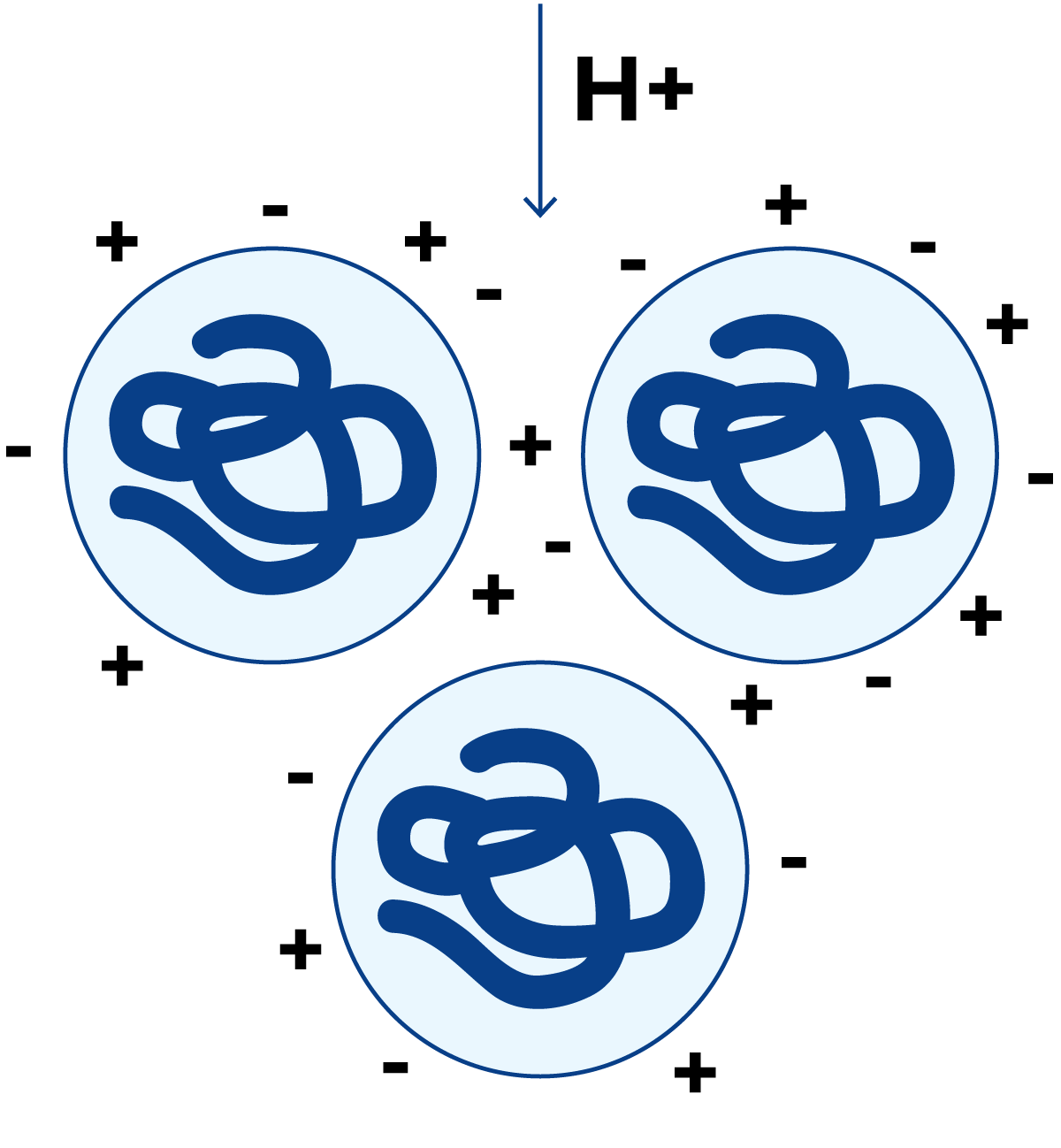
图2.26 pH≈4.6(等电点)时的蛋白质分子
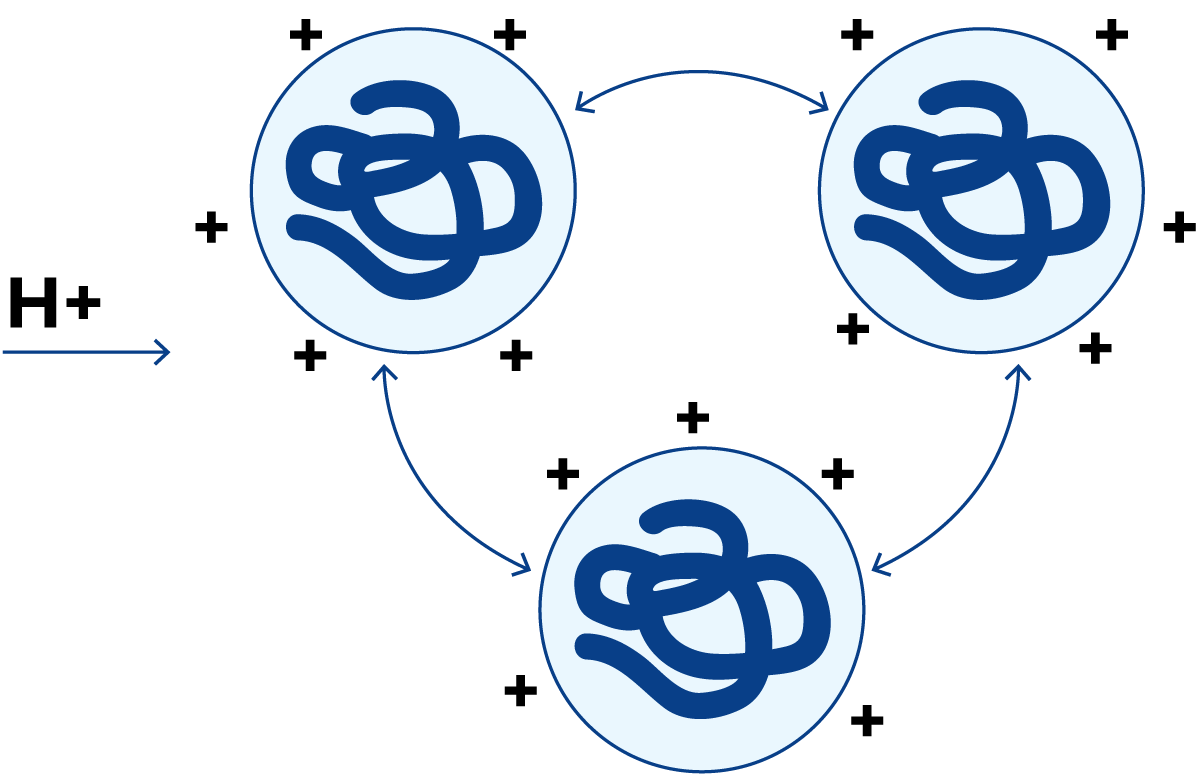
图2.27 pH≈1时的蛋白质分子
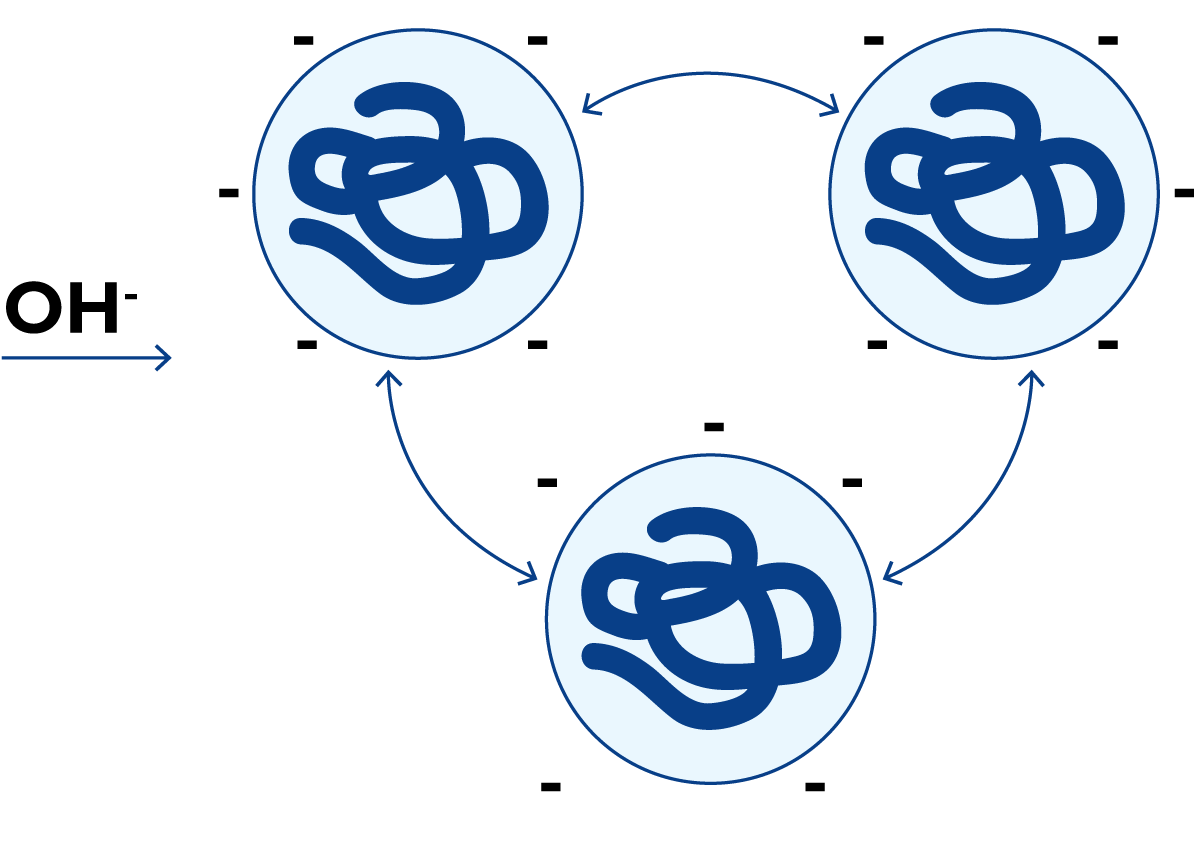
图2.28 pH≈14时的蛋白质分子
在牛奶的正常pH值(≈6.6)条件下,蛋白质分子携带净负电荷(图2.25)。由于相同电荷相互排斥,蛋白质分子保持分散状态。
当添加氢离子时(图2.26),氢离子被蛋白质分子吸附。当达到正、负电荷量相等的pH值时——即侧链上NH3+基团与COO-基团数量相等——蛋白质的净总电荷为零。此时蛋白质分子不再相互排斥,相反,一个分子上的正电荷会与相邻分子上的负电荷结合,形成较大的蛋白质聚集体,最终导致蛋白质从溶液中沉淀析出。此时的pH值称为蛋白质的等电点。
当存在过量氢离子时,蛋白质分子获得净正电荷,如图2.27所示。此时蛋白质分子再次相互排斥,并保持溶液状态。
另一方面,如果添加强碱溶液(NaOH),则所有蛋白质分子都将携带负电荷并溶解。
乳蛋白的分类
牛奶中存在数百种蛋白质,其中大多数含量甚微。根据蛋白质的理化特性及其生理功效,可有各种不同的分类方式。传统的酪蛋白、白蛋白及球蛋白分组体系已被更科学的分类系统所取代。表2.5所示为按现代分类系统分类的乳蛋白简表。为简便起见,未列入微量蛋白质。
乳清蛋白这一名称常用于代表乳浆蛋白,但这一名称应特指干酪加工过程中所产生的乳清中的蛋白质。除乳浆蛋白外,乳清蛋白还包含酪蛋白分子片段。部分乳浆蛋白在乳清中的浓度低于原乳,这一现象是干酪制作前,巴氏杀菌处理使蛋白质发生热变性所致。牛奶中的蛋白质可根据其行为特性与存在形式的显著差异,分为以下三大类:酪蛋白:易于以多种形式从牛奶中沉淀;乳浆蛋白:通常稳定存在于溶液中;脂肪球膜蛋白:顾名思义,附着于脂肪球表面,仅在搅打奶油制作黄油等机械处理下才会被剥离。
酪蛋白
酪蛋白是多种组分的混合物(见表2.5),作为牛奶中的主导蛋白类别,约占乳蛋白总量的五分之四。酪蛋白主要包含四个亚型:αs1-酪蛋白、αs2-酪蛋白、κ-酪蛋白和β-酪蛋白,这些亚型均具有异质性,并由多种基因变异体组成。蛋白质的基因变异体间仅存在少数氨基酸差异。酪蛋白通过自组装,形成大型簇状结构——胶束。每个胶束由数百至数千个独立酪蛋白分子构成,大小从50nm到500nm不等。由于胶束尺寸处于胶体范围内,因此能够散射光线,脱脂乳的白色外观主要源于酪蛋白胶束的光散射效应。
酪蛋白胶束
酪蛋白胶束对牛奶性质有重要影响:它们在很大程度上决定了乳制品在加热和贮存过程中的物理稳定性,是干酪制作工艺的核心要素,并主导发酵乳和浓缩乳制品的流变学特性。
酪蛋白胶束是致密的聚合体,其内部通过磷酸钙形成连接桥键,构建出开放的多孔结构。若通过酸化、添加EDTA或柠檬酸盐等方式去除磷酸钙(CCP-胶体磷酸钙),将导致胶束结构解体。当pH值高于9时,酪蛋白胶束同样会发生解体。
关于其内部结构,长期存在学术争议,迄今尚未完全阐明。学界主要提出了三种理论模型:纳米簇模型、双结合模型及亚胶束模型。但学术界对其以下特征已形成共识:胶束大致呈球形颗粒,平均直径约150nm,但尺寸分布范围较广 。αs-酪蛋白和β-酪蛋白主要集中于胶束中部,κ-酪蛋白则主要集中于胶束表面。胶束周围有一层“毛绒层”,主要由κ-酪蛋白C末端组成,突出胶束表面5-10nm。突出的κ-酪蛋白链具有亲水性并带负电荷,对胶束的空间稳定性起主要作用。若通过添加乙醇或凝乳酶诱导水解等方法去除该毛绒层,胶束的胶体稳定性将被改变,导致其聚集或沉淀。学界普遍认为胶束内存在直径约3nm的磷酸钙“纳米簇”,其囊括了胶束中大部分磷酸盐和钙。维持胶束结构的作用力主要包括:蛋白质基团之间的疏水作用力、纳米簇肽链之间的交联以及离子键。

图2.29 酪蛋白胶束的纳米簇模型*

图2.30 双结合模型*
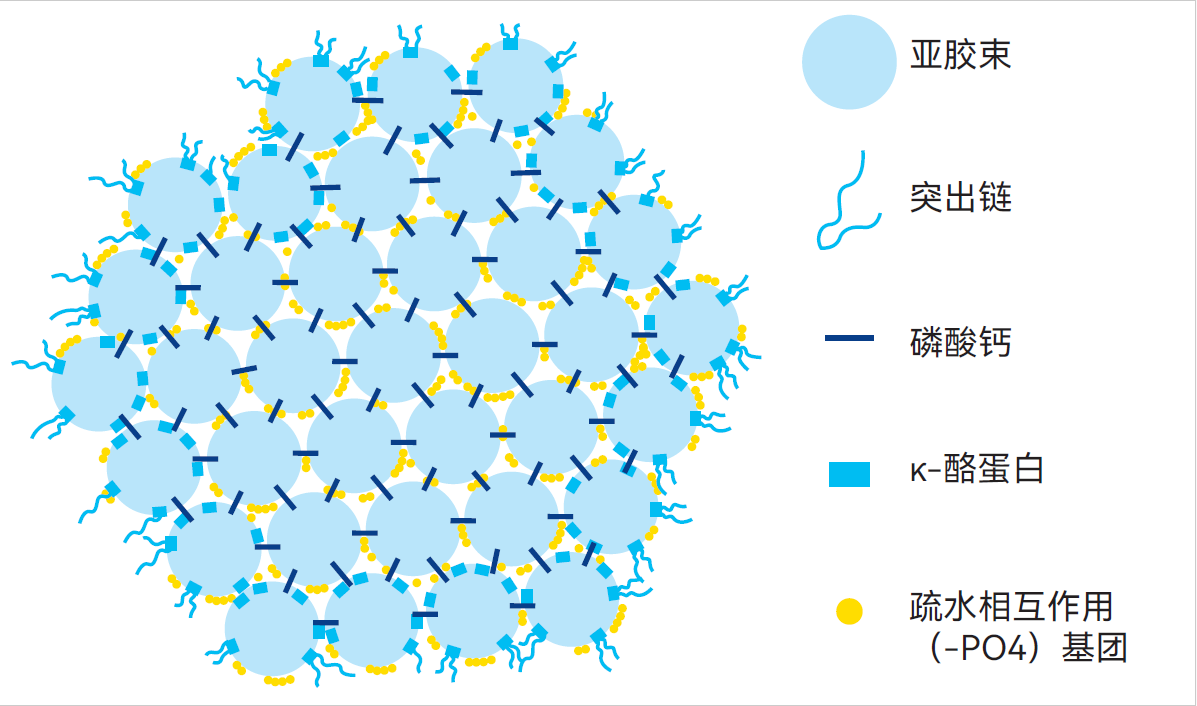
图2.31 酪蛋白胶束的亚胶束模型*
*参考资料:根据Slattery与Evard(1973年)、Schmidt(1982年)及Walstra(1990年)提出的酪蛋白胶束模型摘要重新绘制。依据:Rollema(1992年),Rollema H.S.(1992年)《酪蛋白结合与胶束形成》第63-111页。爱思唯尔科学文献出版社。
纳米簇模型(图2.29,Holt 1992年,De Kruif& Holt 2003年)认为,酪蛋白胶束是由柔性酪蛋白分子组成的缠结网络,通过磷酸钙纳米簇连接形成凝胶状结构。
双结合模型(图2.30)由Horne(1998年)提出,他认为,酪蛋白分子间的疏水相互作用与胶体磷酸钙的交联作用共同维持胶束结构平衡。
亚胶束模型(Morr,1967年;Slattery和Evard,1973年;Walstra,1999年)认为,酪蛋白胶束由直径更小(约10-15nm)的胶束,即亚胶束通过磷酸钙簇连接构成,见图2.31。
酪蛋白胶束结构并非固定不变,而是处于动态平衡状态。胶束与其环境持续进行组分交换,并能响应胶束环境、温度、pH值和压力的变化。
若胶束表面突出的κ-酪蛋白亲水链末端被切割(例如,被凝乳酶切割),胶束将丧失溶解性,开始聚集形成酪蛋白凝乳。完整胶束因携带过剩负电荷而相互排斥。κ-酪蛋白亲水位点结合的水分子是该平衡体系的重要组成部分。若亲水位点被移除,水分子将开始脱离胶束结构,此时,吸引力得以发挥作用,生成新的化学键:其一是钙参与的离子键,二是疏水键。这些键合会加速水分排出,最终使结构坍缩形成致密凝乳。低温同样会对胶束产生不利影响。此时,β-酪蛋白链开始解离,CCP也会脱离胶束结构,以胶体形式存在并溶解于溶液中。对该现象的解释为:β-酪蛋白虽为疏水性最强的酪蛋白类型,但在低温环境下,其疏水作用会减弱。
胶束会表现出解离趋势,且酪蛋白胶束的溶胀度增加。CCP的流失导致单个酪蛋白分子间的吸引力减弱。这些变化会延长凝乳时间并形成质地偏软的凝乳,从而降低乳对干酪制作的适用性。
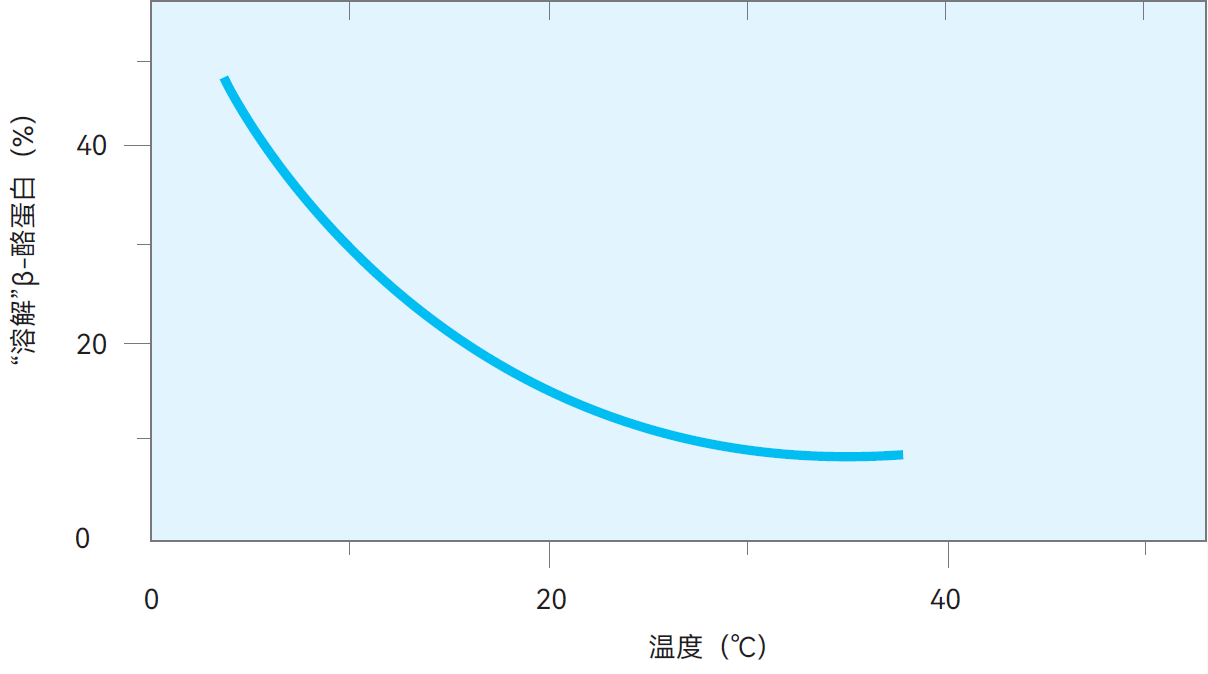
图2.32 牛奶在不同温度下储存约24小时后,未与胶束结合的β-酪蛋白占比
参考资料:根据Walstra、Wouters 和Geurts 2006年著《乳品科学与技术》重新绘制
有两种方式可实现酪蛋白颗粒絮凝和凝固:
- 酸沉淀法
- 酶沉淀法
注意:若向已形成的凝块中添加过量酸,酪蛋白将重新溶解,与酸结合形成盐。若使用盐酸,溶液中将形成部分电离的酪蛋白盐酸盐。
脱离胶束的β-酪蛋白更易被牛奶中多种蛋白酶水解。其水解产物为γ -酪蛋白和䏡蛋白胨,这意味干酪产量降低,因为䏡蛋白胨组分将流失于乳清中。β-酪蛋白的分解也可能形成苦味肽,使乳制品产生异味。不过这些变化过程较为缓慢,在5℃条件下,大约需要24小时才能完成。图2.32显示了β-酪蛋白在24小时储存期内脱离胶束的大致比例(%)。随后将冷藏的原奶或巴氏杀菌奶加热至62-65℃并保持约20秒,β-酪蛋白和磷酸钙将部分回归胶束,从而至少部分恢复乳的原始特性。
温度升高时,胶束会稍微收缩,同时CCP含量增加。当乳浆蛋白存在时,它们会在热变性过程中与酪蛋白胶束结合,且主要附着于胶束表面。β-乳球蛋白与κ-酪蛋白在热处理中的结合即为典型实例。此类结合在冷却后大多不可逆。
酪蛋白的沉淀
酪蛋白的特性之一是其沉淀能力。由于酪蛋白分子及其形成的胶束性质复杂,因此多种不同物质均可引发酪蛋白沉淀。需注意的是,胶束状酪蛋白和非胶束状酪蛋白(如酪蛋白酸钠)的最佳沉淀条件存在显著差异。下面主要介绍胶束状酪蛋白的沉淀。
酸沉淀法
向牛奶中添加酸或让产酸菌在牛奶中生长均会导致pH下降,这将在两方面改变酪蛋白胶束的环境,过程如图2.33所示。首先,酪蛋白胶束中的胶体羟基磷酸钙会溶解并形成离子钙,离子钙会渗透到胶束结构中,并在内部形成牢固的钙桥。其次,溶液的pH值将接近各种酪蛋白的等电点。
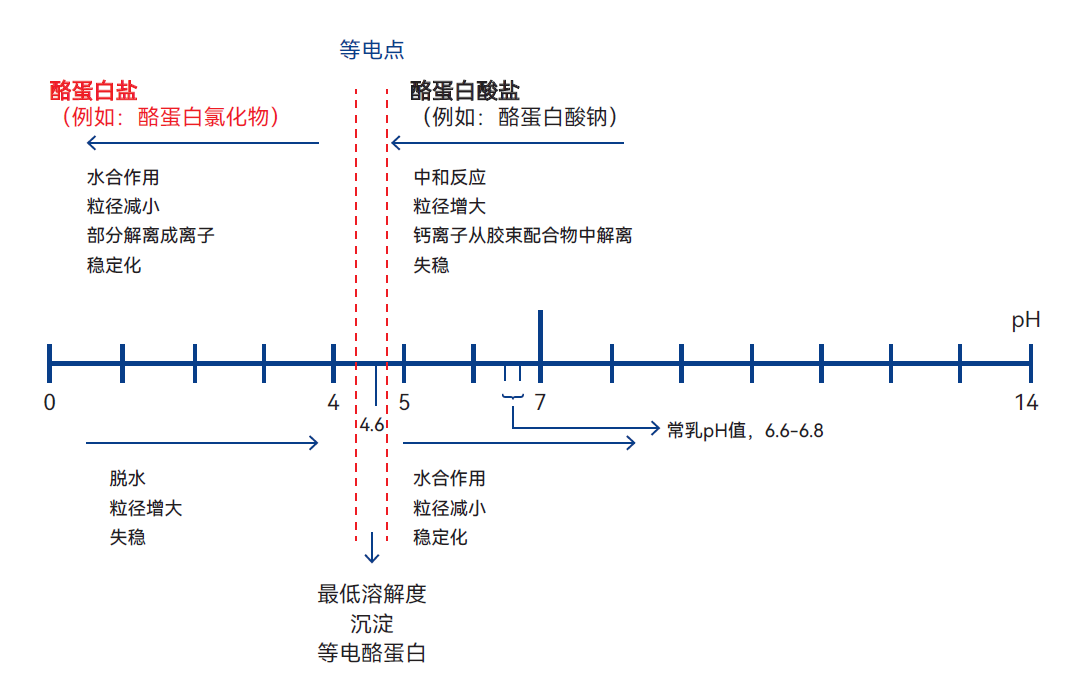
图2.33 酪蛋白受酸碱影响的三个简化阶段
这两种作用机制均会引发胶束内部变化:始于胶束通过聚集作用生长,最终形成致密程度不一的凝块。根据最终pH值的不同,此凝块或含盐形式的酪蛋白,或含等电点状态的酪蛋白,或两者兼而有之。
酪蛋白各组分的等电点取决于溶液中存在的其他离子种类。特定条件下pH的理论值为5.1至5.3。而在与牛奶环境类似的盐溶液中,酪蛋白实现最佳沉淀的pH值范围为4.5至4.9。从牛奶中沉淀酪蛋白的实际应用pH值为4.6。
若向沉淀后的等电点酪蛋白中加入过量氢氧化钠,重新溶解的酪蛋白会转化为酪蛋白酸钠,且部分会解离为离子形态。发酵乳制品的pH值通常在3.9-4.5之间,处于等电点的酸性一侧。在脱脂乳中添加硫酸或盐酸生产酪蛋白时,所选的pH值往往也为4.6。
酶沉淀法
形成k-酪蛋白的氨基酸长链共有169个氨基酸。从酶学角度看,第105位(苯丙氨酸)和第106位(甲硫氨酸)之间的肽键易被多种蛋白水解酶作用。
一些蛋白水解酶会攻击此键位,并将长链切断。可溶性氨基末端包含第106至169位氨基酸,以极性氨基酸和碳水化合物为主,赋予该序列亲水性。κ-酪蛋白分子的这一部分称为“糖巨肽”(GMP),在干酪生产中这一部分释于乳清中。
κ-酪蛋白的剩余部分(第1至105位氨基酸)不溶于水,且与αs-酪蛋白、β-酪蛋白共同生成凝乳,这一部分称为副k-酪蛋白。
凝乳的形成源于亲水性糖巨肽的突然移除导致的分子间作用力失衡:疏水位点间开始键合,且随着胶束中水分子逸出,钙桥进一步增强该过程。这一过程即通常所说的凝乳与脱水收缩阶段。
κ-酪蛋白分子中105-106键的断裂通常称为凝乳酶作用的第一阶段,而凝乳与脱水收缩阶段则称为第二阶段。凝乳酶作用还存在第三阶段,即凝乳酶以更广泛的方式攻击酪蛋白组分,该阶段发生于干酪的成熟期。
这三个阶段的持续时间主要受pH值和温度调控。另外,第二阶段还受到钙离子浓度,以及胶束表面是否附着变性乳浆蛋白这两个因素的强烈影响。
乳清蛋白
乳清蛋白是乳浆蛋白的常用名称。若通过沉淀法(如添加无机酸)从脱脂乳中去除酪蛋白,溶液中残留的一组蛋白质称为乳浆蛋白。未经热变性,即使在其等电点,乳浆蛋白也不会沉淀。
乳受热时,部分乳清蛋白变性并与酪蛋白形成复合物,这会降低酪蛋白被凝乳酶作用及结合钙的能力。经高温处理的牛奶所形成的凝乳不会像普通制干酪凝块那样释放乳清,因其分子内和分子间的酪蛋白桥键数量较少。
一般而言,乳清蛋白(尤其是α-乳白蛋白)具有极高的营养价值,其氨基酸组成非常接近生物学最优标准。乳清蛋白衍生物在食品工业中应用广泛。
α-乳白蛋白
β-乳球蛋白
血清白蛋白
免疫球蛋白
α-乳白蛋白
该蛋白可视为典型的乳清蛋白,存在于一切哺乳动物的乳汁中,并且在乳腺乳糖合成的过程中起着重要作用。
β-乳球蛋白
该蛋白仅存在于有蹄类动物中,是牛奶中主要的乳清蛋白组分。当牛奶加热超过60℃时,其变性过程启动,其中β-乳球蛋白含硫氨基酸的活性发挥关键作用。β-乳球蛋白分子之间、β-乳球蛋白与κ-酪蛋白分子之间、α-乳球蛋白与α-乳白蛋白之间开始形成硫桥。在高温下,含硫化合物(如硫化氢)被逐渐释放出来,这些物质是热处理牛奶产生“蒸煮味”的主要原因。
免疫球蛋白及相关微量蛋白
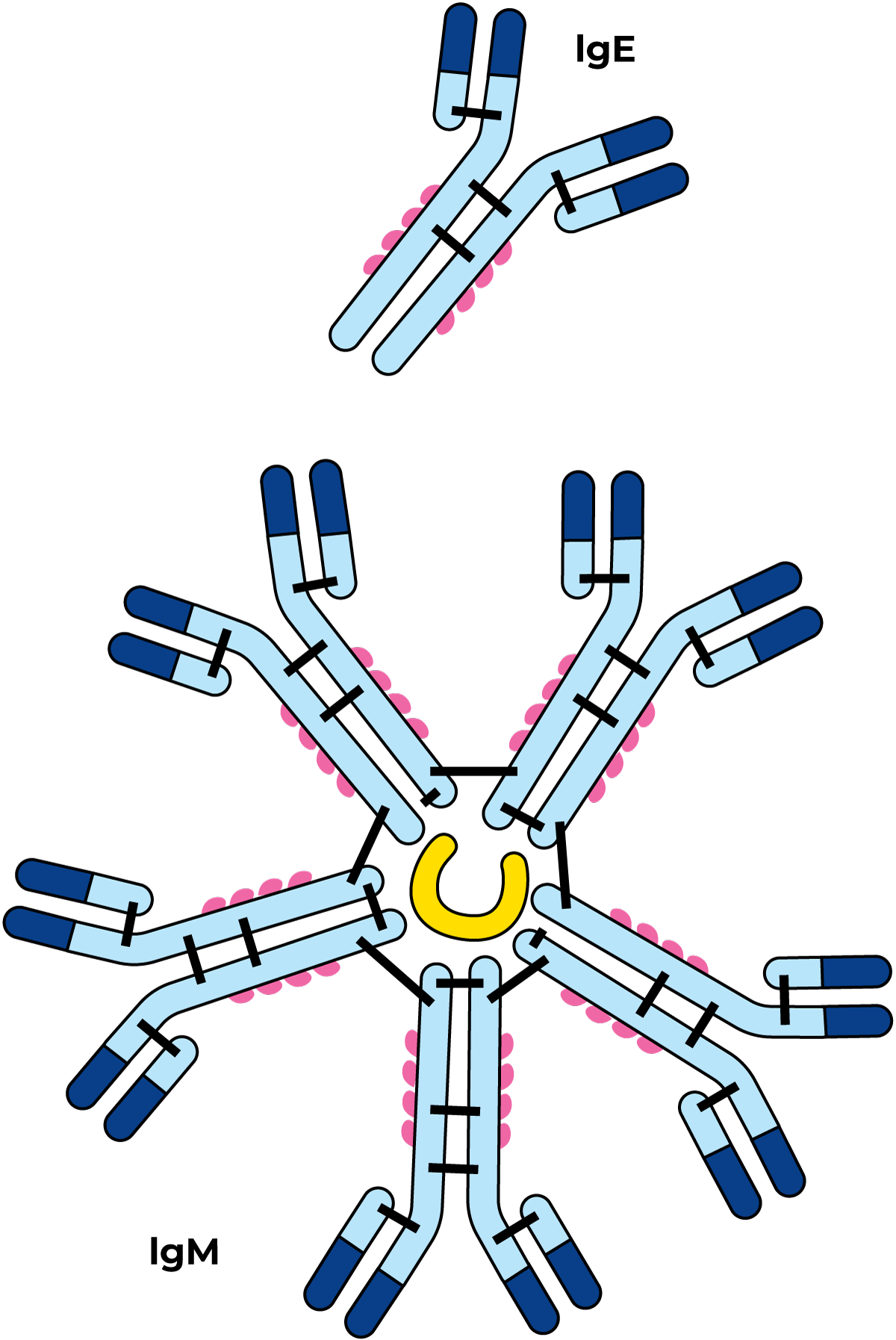
图2.34 两种免疫球蛋白的示意图
参考资料:P.F. Fox和P.L.H. McSweeney,《乳品化学与生物化学》,1998年
这类蛋白质具有高度异质性,且其中仅有少数成员被深入研究(图2.34)。免疫球蛋白是响应特定抗原刺激而合成的抗体。它们主要存在于血液中,在牛奶中的含量较低,但部分免疫球蛋白在初乳和人乳中的含量较高。免疫球蛋白还能针对细菌、病毒乃至脂肪球等“颗粒”发挥作用,使其发生絮凝,该反应被称为凝集反应。通过这种方式,细菌也可与脂肪球发生絮凝,并最终富集在奶油层中。当微生物发生絮凝时,其生长与活动会受到显著抑制。凝集反应对特定抗原具有特异性。不过,部分凝集现象是非特异性的,尤其是在37℃以下的低温乳中发生的所谓“冷沉淀-凝集反应”。这类反应所涉及的蛋白质称为冷球蛋白。凝集素经热处理后失活,其絮凝颗粒的能力随之消失,因此,巴氏杀菌奶中不会发生凝集现象。
未来有望通过商业规模化生产,从乳浆或乳清中分离出多种重要物质。乳铁蛋白与乳过氧化物酶便是两类可能应用于医药及食品工业的物质,目前已能通过商业工艺从乳清中实现分离提取。乳铁蛋白还是一种细菌抑制剂,可抑制包括嗜热脂肪芽孢杆菌和枯草芽孢杆菌在内的细菌,其抑菌机制是通过去除细菌血清中的铁实现的。
膜蛋白
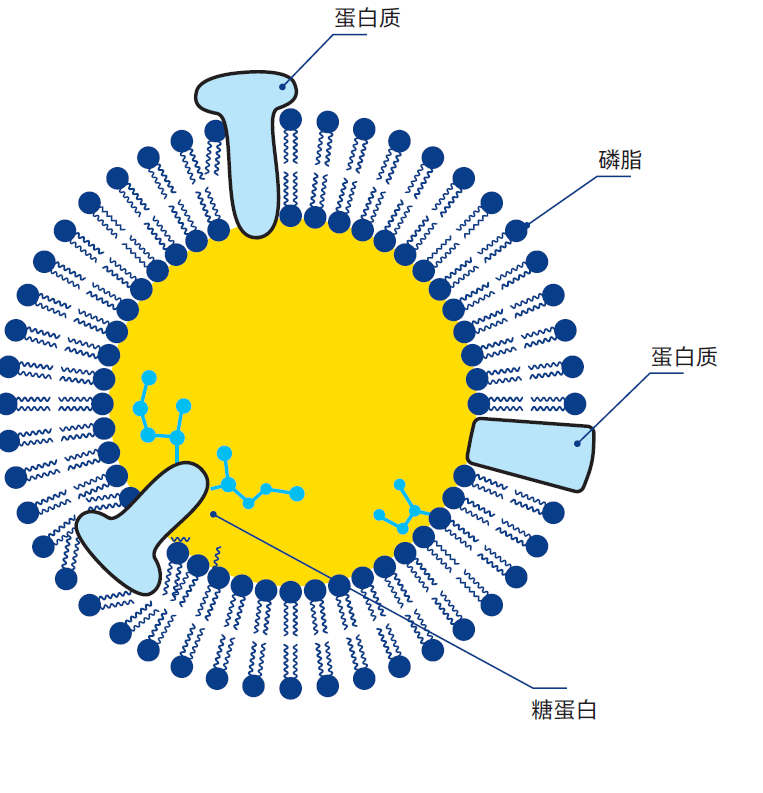
图2.35 膜蛋白覆盖脂肪球表面。
- 磷脂
- 蛋白质
- 糖蛋白
膜蛋白是在脂肪球周围形成保护层以稳定乳液的一类蛋白质(图2.35)。其稠度不一,一些膜蛋白柔软呈果冻状,而另一些则坚韧结实。一些膜蛋白含有脂类残基,称为脂蛋白。这些蛋白的脂质与疏水氨基酸使分子将疏水位点朝向脂肪表面,而疏水性较低部分则朝向水。
弱疏水性膜蛋白以类似方式作用于这些蛋白层,在脂肪表面与水之间形成一个疏水性梯度。这种膜中的疏水性梯度使其成为吸附各种疏水性分子的理想场所。尤其是磷脂和脂肪酶,会被吸附在膜结构内部。只要结构完好无损,酶与其底物之间就不会发生反应;但一旦结构遭到破坏,酶便有机会与底物结合并开始反应。
酶促反应的一个典型例子是:当牛奶在低温下通过故障泵输送,或低温奶经均质处理后未立即进行巴氏杀菌时,便会发生脂肪酸的脂解释放反应。该反应产生的脂肪酸及其他产物,会使产品带有“酸败”味。
变性蛋白质
只要蛋白质所处环境的温度和pH值在其耐受范围内,它们便能保持生物功能。但若被加热至超过特定最高温度,其结构便会改变,此过程称为变性(图2.36)。蛋白质暴露于酸、碱、辐射或高压环境下也会发生相同变化。蛋白质变性后将失去其原有溶解度。

图2.36 部分乳清蛋白质的天然状态(左)和变性状态
蛋白质变性后,其生物活性也会随之终止。酶作为一类催化反应的蛋白质,变性后即丧失催化能力,原因是分子内某些键断裂导致蛋白质结构改变。变性较弱时,蛋白质尚可恢复到其最初状态,恢复生理功能。
但在多数情况下变性是不可逆的,例如煮熟的鸡蛋无法恢复至生蛋状态。
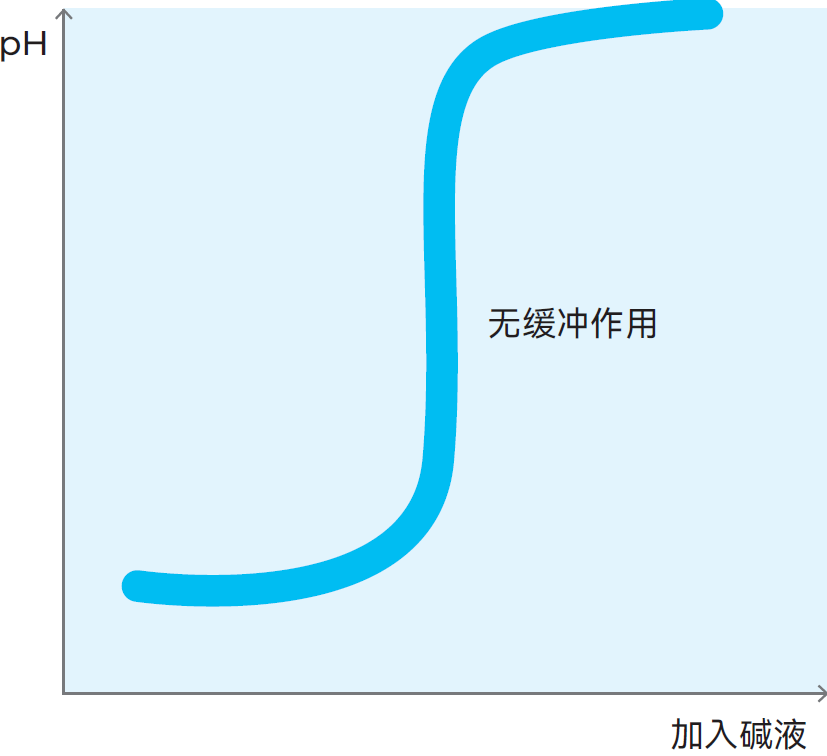
图2.37 若向酸中添加碱,溶液的pH值会立即上升—— 无缓冲作用
牛奶是一种缓冲溶液
牛奶中所含很多物质既能呈现弱酸性,也能呈现弱碱性。例如乳酸、柠檬酸、磷酸及其相应的乳酸盐、柠檬酸盐和磷酸盐。化学上将这些体系称为缓冲溶液,因为在一定范围内,当加入酸液或碱液时,溶液的pH值可以维持不变。这一现象可以通过蛋白质的特性来解释。
当牛奶被酸化时,会添加大量氢离子(H+)。这些离子几乎全部与氨基酸侧链中的氨基结合,形成NH3+离子。但是,由于游离氢离子浓度的增加非常有限,因此pH值几乎不会受到影响。
向牛奶中添加碱时,侧链-COOH基团中的氢离子(H+)被释放,形成COO-基团。因此,pH值实际上保持不变,见图2.38。添加的碱越多,释放的氢离子数量就越多。

图2.38 向牛奶中添加碱时,pH值变化非常缓慢——这表明牛奶中存在显著的缓冲作用
牛奶中一些其它成份也具有类似结合或释出离子的能力,所以当加入酸或碱时,牛奶的pH值改变很缓慢。
然而,在因长期高温储存而已呈酸性的牛奶中,其几乎所有的缓冲容量已被耗尽。此时,仅需添加少量酸即可改变pH值。
牛奶中的酶
酶是一类能够触发化学反应并影响反应进程和速度的蛋白质。酶在反应过程中不会被消耗,因此有时被称为生物催化剂。酶的作用机制如图2.39所示。
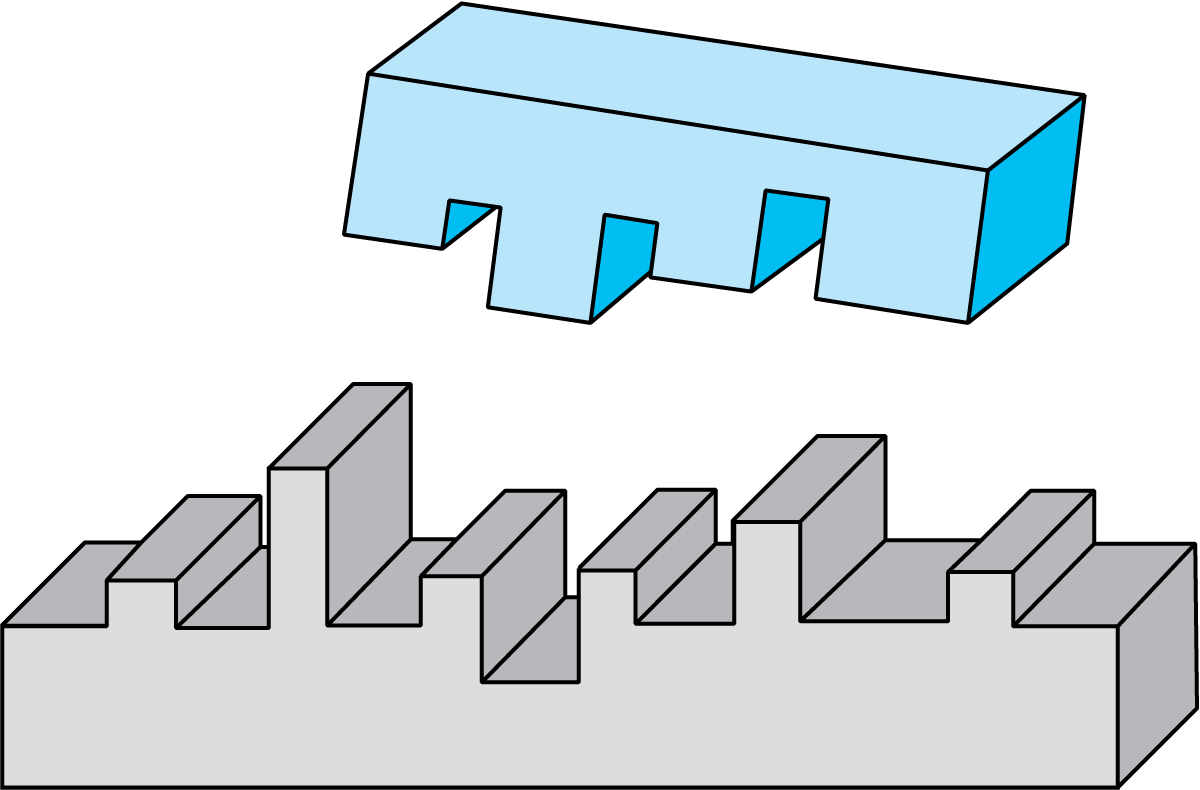
图2.39 一种酶只能作用于特定分子的特定键位
酶进入到分子链的特定位点,削弱化学键。
分子断裂,此时酶游离出来,并以同样的方式结合和作用其他分子。
酶的作用具有特异性——每一种酶只对一种反应起催化作用。影响酶反应的两个重要因素是温度和pH值。一般来说,酶在25-50℃的最适温度范围内活性最高。
当温度超过最适范围后,其活性下降,并在50-120℃之间的某一温度点完全丧失活性。在此温度范围内,酶会发生不同程度的变性(失活)。不同酶类的失活温度存在差异——这一特性已被广泛应用于判定牛奶的巴氏杀菌温度。酶也具有其最适pH值范围:有些酶在酸性溶液中活性最佳,另一些则在碱性环境下功能最强。牛奶中的酶有两个来源,一是母牛乳腺,二是细菌。前者是牛奶的正常成分,被称为“原生酶”。后者,即细菌酶,其种类与含量随菌群性质及规模变化而有所不同。牛奶中的多种酶还被用于乳品的质量检测与控制,其中较为重要的有过氧化物酶、过氧化氢酶、磷酸酶及脂肪酶。
过氧化物酶
过氧化物酶能够将过氧化氢(H2O2)中的氧原子转移至其他易氧化物质。将牛奶加热至80°C保持数秒,该酶即可失活。利用此性质,根据牛奶中是否存在过氧化物酶即可判断巴氏杀菌温度是否到80°C以上。这一检测称为斯托奇过氧化物酶检测。
过氧化氢酶
过氧化氢酶可将过氧化氢分解为水和游离氧。通过测定牛奶中酶释放出的氧总量,可估算牛奶中过氧化氢酶的含量,从而判定牛奶是否来源于健康乳房。患病乳房分泌的乳中含有高浓度过氧化氢酶,而健康乳房分泌的新鲜乳中仅含微量过氧化氢酶。然而有很多种细菌可以产生过氧化氢酶。在75℃下加热60秒,即可破坏过氧化氢酶。
磷酸酶
磷酸酶具有将特定磷酸酯分解为磷酸和相应醇类的特性。可通过向牛奶中添加磷酸酯和一种试剂来检测是否存在磷酸酶,该试剂与游离醇反应时会发生颜色变化。假如颜色变化,则证明牛奶中含有磷酸酶。常规的巴氏杀菌(72℃,保持15~20秒)可破坏磷酸酶,因此可通过磷酸酶检测来确定是否实际达到巴氏杀菌温度。乳品加工业常规采用的检测方法称为“Scharer磷酸酶检测”。
该测试最好在热处理结束后立即进行。如果不能立即进行,则需将牛奶冷却至5℃以下,并在检测之前一直保持此温度。化验工作应在同一天内完成,否则可能出现所谓的“酶复活”现象——即已失活的酶重新恢复活性,导致检测结果呈阳性。在这方面,奶油尤其容易出现这种现象。
脂肪酶
脂肪酶可将脂肪分解为甘油和游离脂肪酸,见图2.40。牛奶和乳制品中的过量游离脂肪酸会产生酸败味。虽然在某些乳中脂肪酶活性可能很强,但多数情况下脂肪酶活性十分微弱。有研究认为,牛奶中脂肪酶的含量在泌乳周期末期会升高。脂肪酶在很大程度上可通过巴氏杀菌失活,但完全失活则需要较高温度。许多微生物均能产生脂肪酶,这可能会引发严重问题,因为这类酶非常耐热。
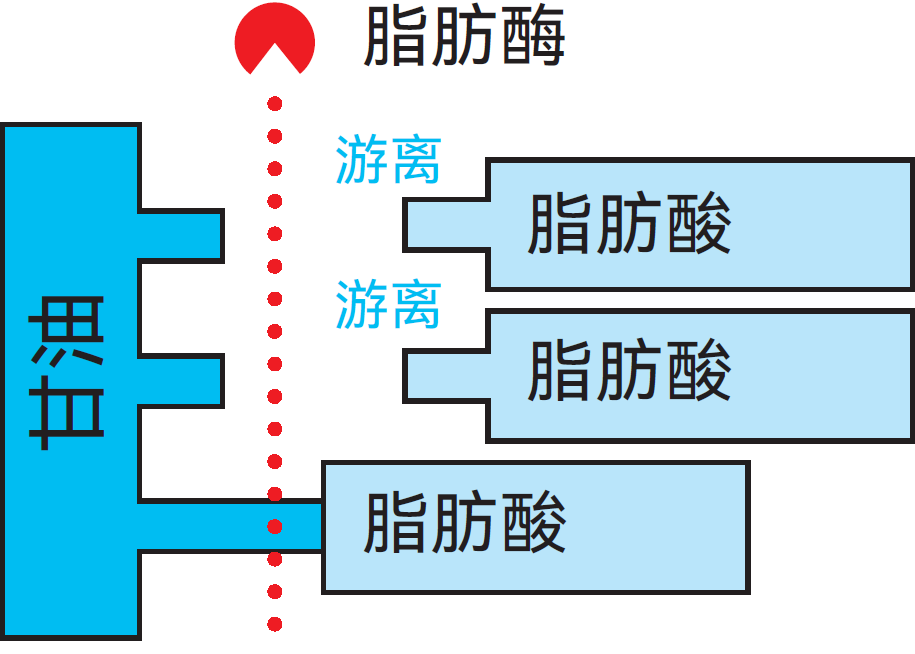
图2.40 脂肪酶分解脂肪示意图
牛奶中的乳糖
乳糖是一种只存在于乳中的糖类,属碳水化合物类有机化合物。碳水化合物是人类膳食中最重要的能量来源,例如面包和土豆中就含有丰富的碳水化合物,能为人体提供充足的营养。它们分解成高能化合物,参与各类生化反应并供给必需能量。碳水化合物也为人体某些重要的化合物合成提供了原料,它们以肌糖原形式存在于肌肉中,以肝糖原形式存在于肝脏中。糖原是一种大分子碳水化合物,淀粉和纤维素也同属此类,这类复合碳水化合物也称为多糖,即由许多葡萄糖分子组成的大分子物质。糖原和淀粉内的分子内有许多支链,而纤维素分子则形成长直链结构。
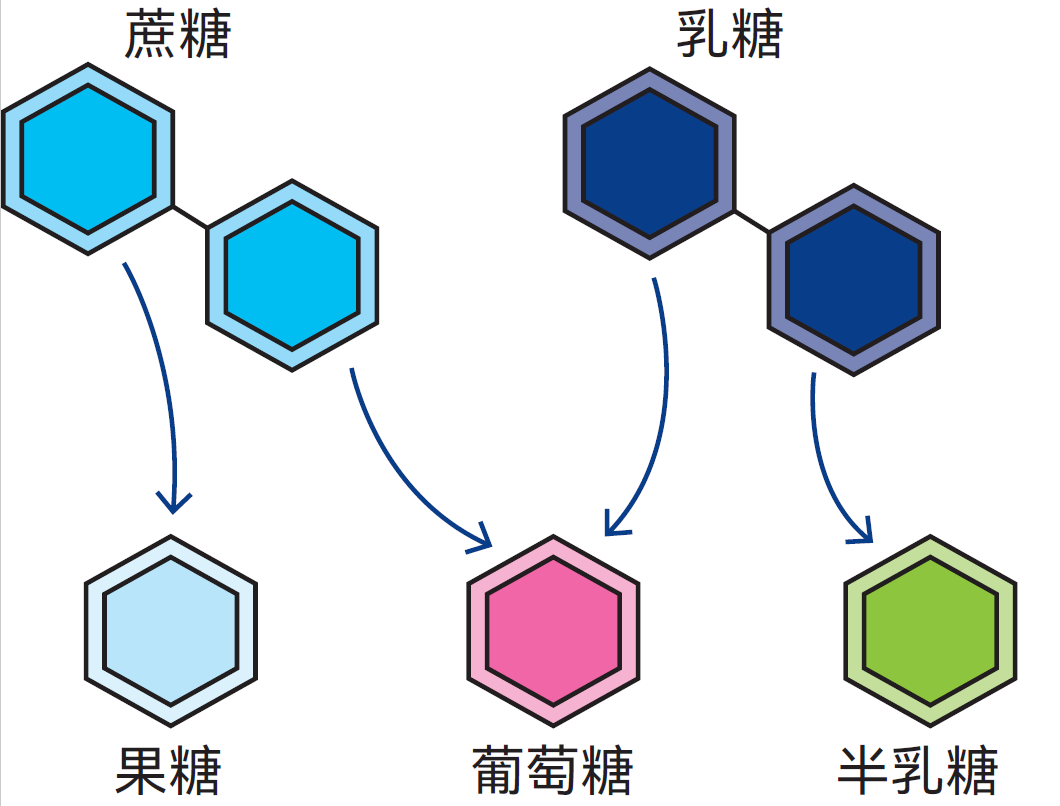
图2.41 乳糖被分解为葡萄糖和半乳糖,蔗糖被分解为果糖和葡萄糖
图2.41展示了几种双糖分子,即由两种糖分子构成的碳水化合物。蔗糖(普通甘蔗糖或甜菜糖)分子由果糖和葡萄糖这两种单糖构成。乳糖(乳中 的糖)同样是双糖,其分子包含葡萄糖和半乳糖这两种单糖。由表2.3可知,牛奶中乳糖含量介于3.6-5.5%之间。图2.42展示了乳酸菌分解乳糖时发生的生化过程。乳糖被转运至细菌细胞内,由胞内酶对其进行作用,将其分解为葡萄糖和半乳糖。随后,乳酸菌中的其它酶类又与葡萄糖和半乳糖反应,通过复杂的中间反应将其转化为以乳酸为主的物质。各种酶在这些反应中依序作用。这就是乳变酸的过程:乳糖被发酵转化为乳酸。牛奶中其它微生物还会产生其它分解产物。
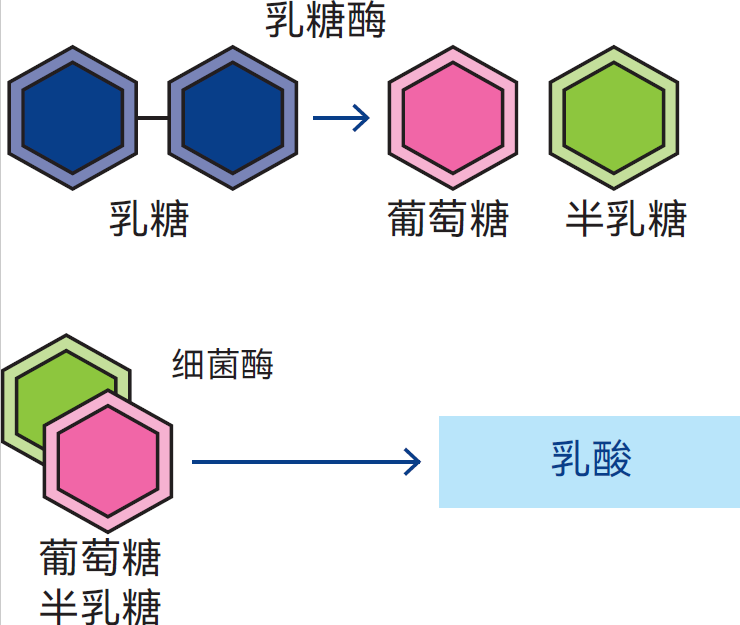
图2.42 酶促作用下的乳糖分解与乳酸形成
如果将牛奶长时间高温加热,牛奶将发生褐变并产生焦糖风味。此过程称为焦糖化,是乳糖与蛋白质发生美拉德反应的结果。
美拉德反应(又称非酶褐变反应,因其无需酶参与即可发生)由热处理引发,并在产品贮藏期间持续进行。该反应动力学直接取决于热负荷和后续贮存温度等因素。乳糖具有水溶性,在牛奶中以分子溶液形式存在。在干酪生产中,大部分的乳糖溶解在乳清中。生产乳清干酪时蒸发乳清可进一步提 高乳糖浓度。乳糖甜度低于其他糖类,例如其甜度比蔗糖低30倍。
牛奶中的维生素
维生素是存在于动植物体内的微量有机物,它们是生命正常生长过程中必不可少的基础物质,但不能由身体合成。维生素的化学组成通常都非常复杂,但是目前大部分维生素的化学成份已为人类所知。维生素采用大写字母命名,有时会附加数字下标,如维生素A、B1和B2。牛奶是维生素的良好来源,牛奶中维生素的含量各有差异。
其中为人所周知的有维生素A、B族维生素、维生素C和D。维生素A和D可溶于脂肪或脂肪溶剂,而其他维生素可溶于水。就脂溶性维生素而言,以维生素A和D最为重要。它们会影响视力和皮肤。由于天然成因,低脂乳制品中这类维生素含量较低。许多国家在低脂乳中添加维生素A和D,使其达到与全脂乳相当的维生素水平。
表2.6列出了一升市售牛奶中的各类维生素含量,以及成人每日维生素需求量的数据。该表说明牛奶是维生素的良好来源。维生素缺乏可能导致各种缺乏症,见表2.7。
牛奶中的矿物质与盐类
牛奶中含有多种矿物质,总浓度低于1%。矿物盐或溶解于乳浆中,或存在于酪蛋白化合物中。最重要的盐类有钙、钠、钾和镁盐,它们分别以磷酸盐、氯化物、柠檬酸盐和酪蛋白酸盐的形式存在。在常乳中,钾盐和钙盐含量最为丰富。盐类的含量并非恒定:泌乳末期时氯化钠含量上升(患乳腺疾病时尤为显著),会使乳产生咸味,此时其他盐类的含量则相应减少。
牛奶中的其它成分
牛奶中始终含有体细胞(白血球或白细胞)。健康乳腺所产乳中体细胞含量较低,但若罹患乳腺疾病,其含量会升高,通常与疾病严重程度成正比。健康乳畜所产乳的体细胞含量通常低于200000个/毫升,但最高可接受上限为400000个/毫升。
牛奶中还含有气体:刚挤出的牛奶中气体含量约占体积的5-6%,但运抵乳品厂时可能升至10%。这些气体主要包括二氧化碳、氮气和氧气。
它们在牛奶中以三种形态存在:
- 溶解于牛奶中
- 与牛奶键合且无法分离
- 分散于牛奶中
分散及溶解的气体在乳品加工中会引发严重问题——若牛奶中气体含量过高,加热时易在加热表面产生结焦现象。
牛奶及其成份的变化
贮存过程中的变化
在贮存期间,牛奶中的脂肪和乳蛋白可能会发生化学变化。这些变化通常包括两种:氧化和脂解。这两种反应的产物可产生异味,主要表现为牛奶与黄油中出现异味。
脂肪氧化
脂肪氧化会产生金属味,并使黄油呈现油腻 味、哈喇味。氧化作用发生在不饱和脂肪酸的双键处,其中卵磷脂的双键最易被氧化。铁盐与铜盐的存在会加速自动氧化进程和金属味的形成,溶解氧的存在以及光照(尤其是直射日光或荧光灯照射)同样会加速该过程。
脂肪氧化可通过三种方式部分抑制:一是利用牛奶中的微生物,二是采用80℃以上温度进行巴氏杀菌,三是添加抗氧化剂(还原剂)。乳酸菌等微生物消耗氧气,还具有还原作用。在低温条件下,这些微生物的活性降低,故而易使乳制品氧化而带有异味。另外,氧气在牛奶中的溶解度随温度下降而升高。高温巴氏杀菌可缓解该问题,因牛奶在加热时会产生含-SH基团的还原性化合物。
金属氧化异味在冬季的发生率高于夏季,这部分归因于较低的环境温度,部分源于奶牛日粮结构的变化。夏季饲料富含维生素A和C,增加了牛奶中还原物质的数量。
在光照和/或重金属离子作用下,脂肪酸还会进一步分解为醛、酮,这些物质会引发异味,如脂肪类乳制品中的酸败味。
蛋白质氧化
当暴露于光线时,蛋氨酸会在核黄素(维生素B2)和抗坏血酸(维生素C)的复杂参与下降解为3-甲硫基丙醛。3-甲硫基丙醛或3-巯基-2-甲基丙醛是“阳光味”(该特殊风味的称谓)的主要来源。由于蛋氨酸并非以游离态存在于牛奶中,而是乳蛋白的组成成分之一,因此,蛋白质的分解必然伴随异味的产生。
导致阳光味产生的因素包括:
- 光照强度(日光和/或人工光源,尤其是荧光灯)。
- 光照持续时间。
- 牛奶的特定性状:均质牛奶比非均质牛奶对光照更敏感。
- 包装材料材质:不透明的包装(如塑料与纸材)在常规条件下能提供有效保护。
另参见第9章关于巴氏杀菌乳品质维护的相关内容。
脂类分解
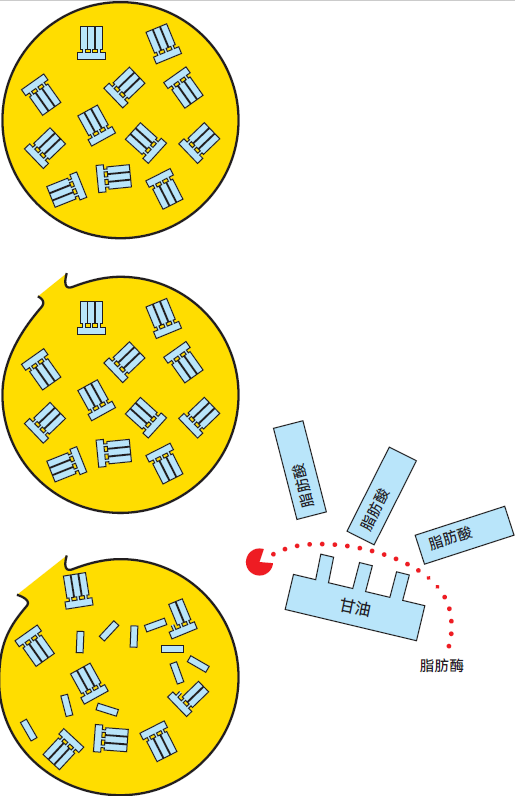
图2.43 当脂肪球膜受损时,脂类分解并释放脂肪酸
脂肪分解为甘油和游离脂肪酸的过程称为脂类分解。由于低分子游离脂肪酸(丁酸和己酸)的存在,脂类分解会产生一种脂肪酸败味。
脂类分解由脂肪酶作用引发,并随贮藏温度升高而加剧。但脂肪酶唯有在脂肪球受损、脂肪暴露后才能发挥作用,见图2.43。只有此时脂肪酶才能作用于脂肪分子并使其水解。在常规乳品加工环节中,脂肪球可能因泵送、搅拌、飞溅等操作而受损。因此,未经巴氏杀菌的牛奶应避免过度搅拌,否则可能引发脂肪酶大面积作用,释放脂肪酸,导致牛奶产生酸败味。必须通过高温巴氏杀菌使脂肪酶失活,以防止其分解脂肪。该处理可完全破坏原生酶。然而,细菌酶的耐热能力较强,即使超高温灭菌处理也不能使其完全失活。(UHT=超高温灭菌处理,即加热至135-150℃或更高温度,持续几秒钟。)
热处理的影响
乳品厂会对牛奶进行热处理,以杀灭任何可能存在的致病微生物。热处理同时会引起牛奶成分的变化:温度越高、受热时间越长,变化越显著。在特定范围内,时间与温度参数可相互协调——短暂高温加热可产生与长时间低温处理相当的效果。因此在热处理时必须考量时间与温度两个参数。
脂肪
在牛奶的主要成分中,脂肪可能受热处理的影响最小。然而热处理确实会引起乳脂肪的变化,尤其体现在物理特性方面。热处理对乳脂肪的主要影响体现在脂肪球的分层上浮方面。
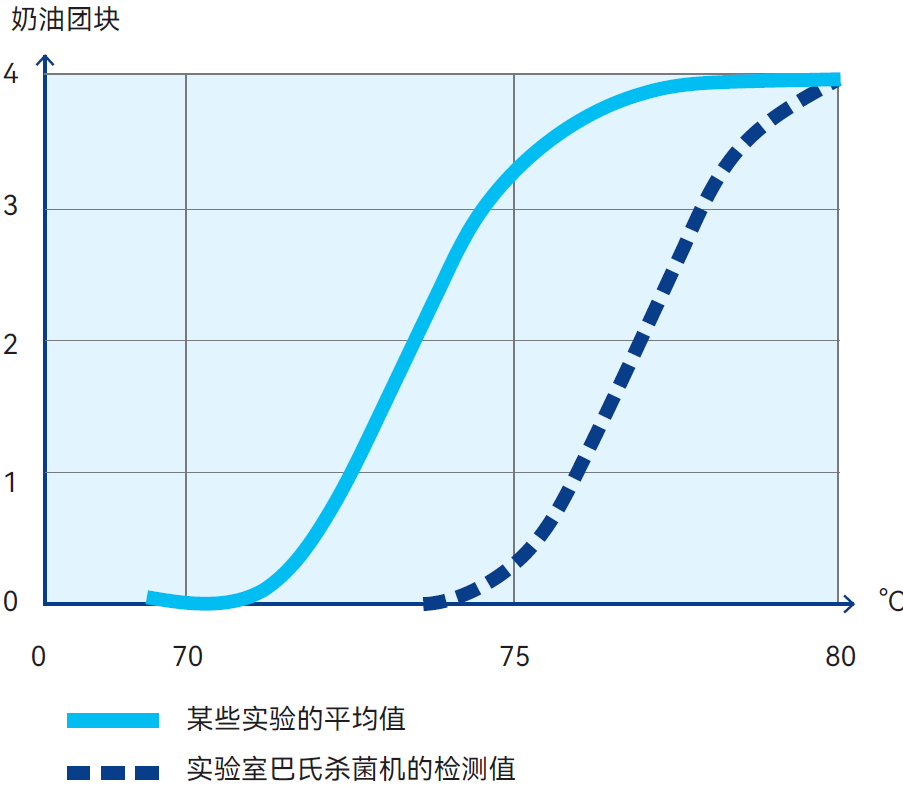
图2.44 牛奶中奶油团块的形成与巴氏杀菌温度的函数关系。
评估标度:0(无影响)到4(固态奶油团块)。所有巴氏杀菌时间都非常短(大约15秒)。参考资料:Thomé 等人
研究表明,将牛奶在70-80℃下进行15秒巴氏杀菌,会形成奶油团块(Thomé等人,Milchwissenschaft 13,115页,1958年),见图2.44。学界已针对该现象探讨过多种理论,但现有研究表明,当脂肪球相互碰撞时,释放出的游离脂肪会将其粘结在一起。为避免形成奶油团块,推荐采用均质处理。
A.Fink与H.G.Kessler(Milchwissenschaft 40.6-7,1985年)的研究表明:含脂率30%的奶油——无论是否经过均质——在加热至105-135℃时,都会有游离脂肪从脂肪球逸出。这被认为是因为脂肪球膜失去稳定性,导致渗透性增加引起的,其结果是可提取的游离脂肪在相互碰撞的脂肪球之间起到粘合剂的作用,并产生稳定的团簇。
当温度超过135℃时,沉积在脂肪球膜上的蛋白质会形成网状结构,使膜层致密化并降低其渗透性。因此,对含脂率较高的乳制品进行超高温灭菌处理时,建议均质操作设在灭菌之后。
蛋白质
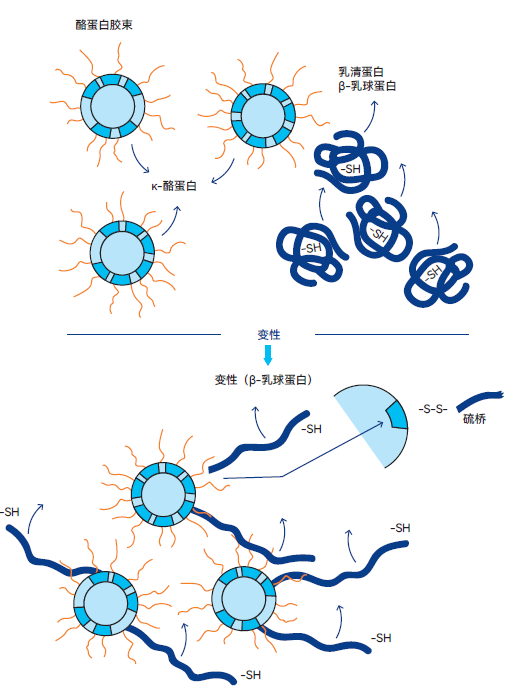
图2.45 变性过程中,k- 酪蛋白粘附在β-乳球蛋白上
酪蛋白是牛奶中主要蛋白质,在正常的pH值、盐类及蛋白质含量范围内,通常不会因加热而变性。
相反,乳清蛋白(特别是约占乳清蛋白总量50%的β-乳球蛋白)对热相当敏感。乳清蛋白在65℃开始变性,加热至90℃维持五分钟则几乎全部变性。
乳清蛋白热变性属于不可逆反应。随机卷曲的蛋白质结构会“展开”,特别是β-乳球蛋白,会通过硫桥与突出的κ-酪蛋白链结合。该高度概括的转化过程如图2.45所示。阻断大部分κ-酪蛋白会削弱牛奶的凝乳能力,因为干酪制作中所用凝乳酶难以从酪蛋白胶束中分离κ-酪蛋白。在恒定保温时间下,巴氏杀菌温度越高,形成的凝块越柔软。这种现象在半硬质和硬质干酪生产中属于不利因素。然而,由于原奶制成的干酪存在致病菌存活和繁殖的风险(特别是单核细胞增生李斯特菌),基于食品安全考量, 推荐对干酪生产所用的原奶进行巴氏杀菌,温度为72℃,时间为15-20秒。虽然原奶也可用于制作干酪,但须接受国家法规的严格管控。
在发酵乳制品(如酸奶)的加工中,将原奶在90-95℃下加热3-5分钟,可促使乳清蛋白质变性并与酪蛋白反应,这一过程有助于减少脱水收缩、改善粘度,从而提升产品品质。牛奶在75℃下加热20-60秒会产生“蒸煮味”,这源于β-乳球蛋白及其他含硫蛋白质(失活脂蛋白)释放的硫化合物。
酶类
加热可使酶失活,但酶的失活温度因种类不同而存在差异。
在冷藏原奶及热处理乳制品的腐败菌群中,假单胞菌属是常常被提及的典型菌群。这类细菌能产生耐热性极强的蛋白水解酶和脂肪酶——即使对牛奶实施巴氏杀菌或超高温灭菌处理,也仅能抑制其部分酶活性。
乳糖
牛奶中的乳糖比在干燥状态下的乳糖更易发生变化。当温度超过100℃时,乳糖与蛋白质会发生反应,呈现褐色。这一系列反应由氨基酸残基的氨基与牛奶碳水化合物的醛基共同参与,称之为美拉德反应(亦称褐变反应)。此反应会导致产品褐变、风味改变、营养价值降低——尤其是作为必需氨基酸之一的赖氨酸含量的降低。
乳果糖是乳糖的差向异构体,在牛奶经热处理后生成(Adachi,1958年)。其含量随热处理强度增加而增加,因此可作为区分巴氏杀菌乳、超高温灭菌奶的指标(Martinez Castro & Olano,1982年;Geier & Klostermeyer,1983年)。
维生素
牛奶是维生素A、D及B族维生素的重要来源。脂溶性维生素具有高度热稳定性,其含量不会因热处理而降低。但牛奶中添加的维生素A相对损耗率较高。
维生素损耗主要涉及维生素C和部分B族维生素。维生素C的损耗通常影响较小(因牛奶非其主要来源),但仍会影响营养价值。维生素C的分解与维生素B12的分解相关联,同时能保护叶酸免于氧化。
维生素的降解不仅与热处理相关,还受最终产品贮藏条件影响。若有效隔绝氧气和光照,可大幅避免贮藏期间的维生素损耗。高氧环境下,维生素C与B9可能会在数日内完全消失。该反应由核黄素(维生素B2)催化,而光照会加速反应进程。长期光照会导致大部分核黄素消失。不同处理方式造成的维生素损耗情况如表2.8所示。
矿物质
磷酸钙的溶解度与温度密切相关。与大多数化合物不同,磷酸钙的溶解度随着温度的升高而降低。这意味着加热会导致胶束中以CCP形式存在的磷酸钙沉淀,而冷却则会增加可溶性磷酸钙的浓度。冷却后,该反应易发生逆转,但经高温加热后,逆转过程变得更加缓慢且不完整。高温下的变化会导致牛奶的酸度升高,pH值下降,如表2.9所述。
pH值的变化机制如下所示:

公式 2.1
当牛奶被浓缩时,pH值的变化会加剧。
牛奶的物理性质
外观
牛奶的不透明性源于脂肪、蛋白质及某些矿物质形成的悬浮颗粒。牛奶的颜色介于白色至淡黄之间,具体取决于脂肪的色泽(胡萝卜素含量)。脱脂乳透明度较高,并稍带蓝色。
密度
牛奶密度通常介于1.028-1.038g/cm³之间,具体取决于其成分组成。15.5℃时的牛奶密度可按下列公式计算:

公式 2.2

示例
渗透压
渗透压取决于分子或颗粒数量,与溶质重量无关。因此,100个粒径为10的分子产生的渗透压是10个粒径为100的分子的10倍。由此可知:在重量相同的情况下,分子越小则渗透压越高。
牛奶由血液经渗透膜分离形成,故二者渗透压相同,即牛奶与血液呈等渗状态。尽管血液在色素、蛋白质等成分上可能存在差异,但其渗透压非常稳定。如表2.10所示,牛奶的总渗透压同样遵循这一规律。
参考资料:《乳业词典》,作者:J. G. Davis
冰点
牛奶的冰点是检测是否掺水的唯一可信的参数。研究发现,单头奶牛所产牛奶的冰点变化范围在-0.51--0.55℃之间。此外需说明,当牛奶经高温处理(超高温处理或灭菌)时,部分磷酸盐的沉淀会导致冰点升高。
内压(即渗透压)也决定了溶液与溶剂(水)之间的冰点差,因此冰点降低值(表2.10中的D值)可作为渗透压的衡量指标。当牛奶成分因生理或病理原因(如泌乳末期或乳腺炎)发生变化时,虽被归类为异常牛奶,但其渗透压及相应的冰点保持恒定。最重要的改变是乳糖含量的降低和氯化物含量的升高。
酸度
溶液的酸度取决于其中氢离子[H+]的浓度。当[H+]和[OH-](氢氧根)离子的浓度相等时,溶液为中性。在中性溶液中,每升溶液中[H+]的数量为1:10000000mol/l或10-7mol/l。pH值表示溶液中的氢离子浓度,在数学上可以定义为氢离子[H+]浓度的负对数。
应用于上面的示例, p H 值为p H = -log10-7=7,这是中性溶液的典型值。当[H+]为1:1000000mol/l或10-6mol/l时,pH值为6,溶液为酸性。因此pH值越小,酸性越强。溶液或制品的pH值标志着它们的即时酸度(真实酸度)。常乳是一种弱酸溶液,在25℃测量温度下,其pH值通常介于6.6至6.8之间,最常见值为6.7。pH值通过pH计进行检测。
滴定酸度
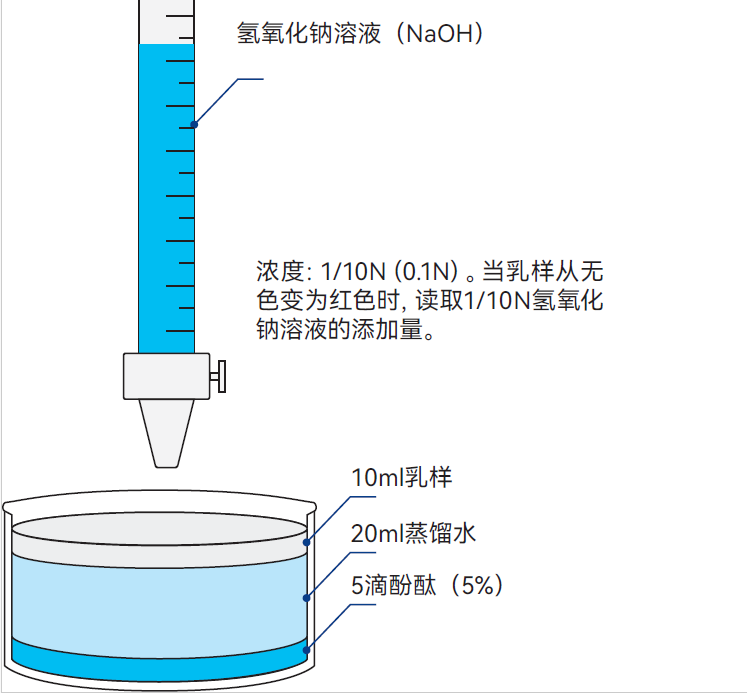
图2.46 以Thörner度(°Th)测定酸度的过程
酸度还可以用滴定酸度来表示。牛奶的滴定酸度是指:将特定量乳样的pH值提升至约8.4(最常用指示剂酚酞由无色变为粉红色的pH值)时,所需特定浓度氢氧根离子(OH-)溶液的量。该检测的本质是测定将pH值从6.7升至8.4所需的碱量。
若牛奶因细菌活动而酸败,则需消耗更多碱液,牛奶的酸度(即滴定值)随之升高。滴定酸度可用多种单位表示,基本上取决于滴定所需氢氧化钠(NaOH)的浓度。
°SH = 即Soxhle Henkle度,取100ml乳样,用1/4N的NaOH进行滴定,以酚酞作指示剂,典型牛奶的°SH值约为7。此方法多被中欧所采用。
°Th = 即Thürner度。取100ml乳样,用2倍蒸馏水稀释,用1/10N的NaOH进行滴定,以酚酞作指示剂,典型牛奶的°Th值约为17。此方法多被瑞典所采用。
°D = 即Dornic度。取100ml乳样,用1/9N的NaOH进行滴定,以酚酞作指示剂,典型牛奶的°D值约为15。此方法多被荷兰和法国所采用。
% l.a. = 即乳酸度。即通过°D值除以100获得。此方法广泛被美国、加拿大、澳大利亚和新西兰所采用。表2.11综合了滴定酸度的各种表达方式。图2.46展示了以Thörner度测定酸度的过程。
滴定10ml乳样需要消耗1.7ml的1/10N NaOH。因此,滴定100ml乳样需要消耗10* 1.7 = 17ml,因此该乳的酸度为17°Th。
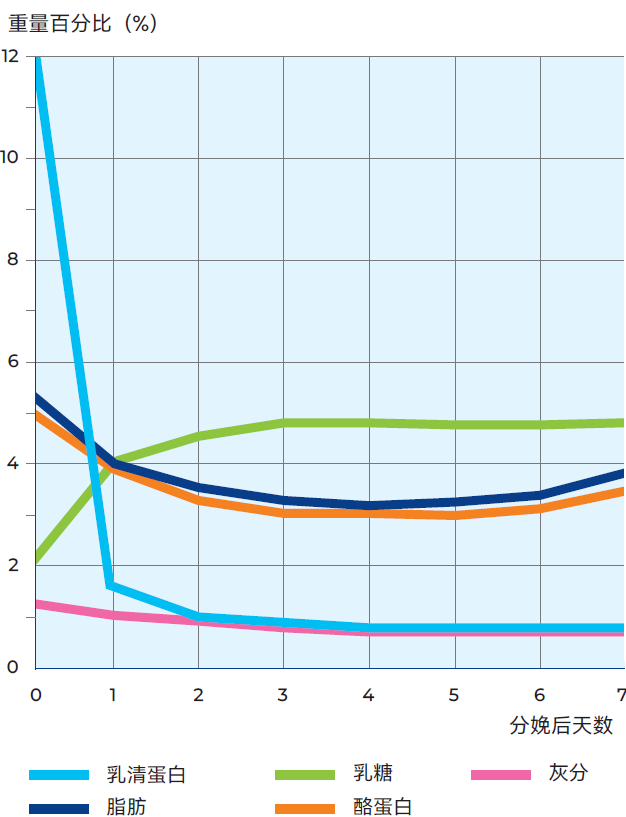
图2.47 奶牛分娩后乳汁成分的变化
初乳
母牛产犊后,最初分泌的乳汁为初乳。其成份和特性与常乳差别很大,一个非常明显的特征是,初乳含大量的乳清蛋白质,其含量高达11%,而常乳中该含量只有0.65%左右, 如图2.47所示。
因此,初乳在加热时易凝固。乳清蛋白中相当大一部分是免疫球蛋白(lg G,在初乳中占主导地位),可保护牛犊在自身免疫系统健全之前免受感染。初乳为浅黄至深棕色,气味特殊并略有咸味,过氧化氢酶和过氧化物酶含量较高。母牛产犊四、五天后,开始分泌出成分正常的牛奶,此时的牛奶可与其它牛奶混合。